
Journal of Plant Biology & Soil Health
Download PDF
Research Article
*Address for Correspondence: Yong-Lak Park, Division of Plant and Soil Sciences, West Virginia University, Morgantown, WV 26506, Tel: 304-293-2882; E-mail: yong-lak.park@mail.wvu.edu
Citation: Gyawaly S, Park YL. Feeding Potential and Prey Acceptance of Podisus maculiventris (Hemiptera: Pentatomidae): Implications for Biological Pest Control.J Plant Biol Soil Health. 2013;1(2): 5.
Copyright © 2013 Gyawaly S, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Biology & Soil Health | ISSN: 2331-8996 | Volume: 1, Issue: 2
Submission: 05 September 2013 | Accepted: 11 October 2013 | Published: 18 October 2013
Feeding Potential and Prey Acceptance of Podisus maculiventris (Hemiptera: Pentatomidae): Implications for Biological Pest Control
Sudan Gyawaly and Yong-Lak Park*
- Division of Plant and Soil Sciences, West Virginia University, Morgantown, WV, 26506, USA
*Address for Correspondence: Yong-Lak Park, Division of Plant and Soil Sciences, West Virginia University, Morgantown, WV 26506, Tel: 304-293-2882; E-mail: yong-lak.park@mail.wvu.edu
Citation: Gyawaly S, Park YL. Feeding Potential and Prey Acceptance of Podisus maculiventris (Hemiptera: Pentatomidae): Implications for Biological Pest Control.J Plant Biol Soil Health. 2013;1(2): 5.
Copyright © 2013 Gyawaly S, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Biology & Soil Health | ISSN: 2331-8996 | Volume: 1, Issue: 2
Submission: 05 September 2013 | Accepted: 11 October 2013 | Published: 18 October 2013
Abstract
Determination of feeding potential and prey acceptance of natural enemies is crucial to assure that natural enemies can control target pest species. This study was conducted to investigate feeding potential and prey acceptance of a generalist predator, Podisus maculiventris (Say) (Hemiptera: Pentatomidae). A series of laboratory experiments were conducted to quantify feeding potential of P. maculiventris by measuring daily feeding amount and to determine prey acceptance by analyzing feeding behavior of P. maculiventris on various types of prey. The result of this study showed that the amount of prey consumption differed significantly (F = 38.48; d.f. = 5, 44; P < 0.05) among stages of P. maculiventris. P. maculiventris attempted to attack Halyomorpha halys (Hemiptera: Pentatomidae), but could not sustain effective feeding due to behavioral and chemical defense of the prey. P. maculiventris showed significantly (P < 0.05) less effective feeding on prey with spines or hairs such as Evergestis rimosalis (Lepidoptera: Crambidae) and Hyphantria cunea (Lepidoptera: Arctiidae). Utilization of P. maculiventris for controlling various types of pests for augmentative biological control is discussed in this article.Keywords
Spined Soldier Bug; Feeding Potential; Prey Acceptance; Generalist Natural EnemyIntroduction
Since the first successful biological control using vedalia beetles to control cottony cushion scale in the United States, many specialist and generalist predators have been used for biological control of insect pests [1]. Specialist predators have a high degree of prey specificity and are considered appropriate to use to reduce a specific pest population below economic injury level [2]. Historically, the view among biological control practitioners was that generalist predators are less important than specialist predators, but recently the importance of generalist predators has been emphasized [3]. Edwards et al . [4] and Chiverton [5] showed that generalist predators in certain manipulated systems could be as important as specialist predators in other systems. These generalist predators include carabids [6], coccinellids [7], neuropterans [8], and predatory hemipterans [9].Major advantages of using generalist predators is their ability to control multiple insect pests and survive on plant material or alternative prey even when their major prey is scarce or absent [8,10]. For example, the multicolored Asian lady beetle Harmonia axyridis (Coleoptera: Coccinellidae) can survive by feeding on pollen and eggs of beetles and moths when its major prey (i.e. aphids) is scarce in the field [11]. Therefore, the presence of alternative prey or food sources in a crop field could improve biological control using generalist predators [12]. Although generalist predators have such advantages they generally have different degrees of acceptance to various types of prey [13]. Prey characteristics, such as host breadth, prey chemistry, and prey behavior and morphology, can play important roles in the acceptance of prey [14]; for example, predatory hemipterans are known to exhibit prey preferences based on differences in prey mobility [12], size [15], and prey species [16]. In addition to prey acceptance, understanding feeding capacity of different stages of generalist predators is crucial to aid mass rearing and release in biological control programs. Because younger predators tend to feed less than mature predators, quantification of feeding potential of younger stages of a predator relative to older ones will help determining the number of predators to release to control pests in the field and deciding the optimal amount of prey needed for mass rearing of the predators.
The spined soldier bug Podisus maculiventris (Hemiptera: Pentatomidae) is a generalist predator feeding on more than 70 species of insects [17], mostly immature stages of insects in the orders of Lepidoptera and Coleoptera [18,19]. P. maculiventris can inhabit various agricultural ecosystems (i.e. crop fields, forests, and grasslands) and plays a key role in the control of major insect pests [17,20]. Specifically, P. maculiventris is an important predator of field crop pests [18] including many larvae of noctuids (Lepidoptera: Noctuidae) and Colorado potato beetles (Coleoptera: Chrysomelidae). In addition, P. maculiventris has high potential for augmentative biological control because it has a high reproduction rate making mass production easier [21].
This study was conducted to investigate the feeding potential and prey preference of P. maculiventris for augmentative biological control. The objectives of this study were determining feeding potential of different P. maculiventris stages/instars for a mass rearing system (Experiment 1) and investigating prey acceptance and efficacy of P. maculiventris as a generalist predator (Experiments 2 and 3).
Materials and Methods
Experimental conditions and insectsThis study was conducted at a constant temperature (22 ± 1.7°C), relative humidity (60 ± 8.5%), and photoperiod (15:9 L:D). All P. maculiventris used in the experiment were obtained from colonies that have been maintained in the entomology laboratory at West Virginia University (Morgantown, WV) by using the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae) as prey. Young T. molitor (third instars) were purchased from New York Worms (Long Island, New York, NY) and reared them in the laboratory to fifth instars and pupae that were used in this study.
Experiment 1: Feeding potential of different P. maculiventris stages
An experiment was conducted to quantify daily feeding amount of P. maculiventris expressed as weight loss of prey. T. molitor was used as prey in this study because it is common prey used for mass rearing of P. maculiventris. All predacious stages of P. maculiventris were used in this study (i.e. all but egg and first instar). One T. molitor pupa was weighed by using an electronic balance (PB 303-S/Fact, Mettler-Toledo Inc., Columbus, OH) and placed in a cup (4-cm diameter) (Solo®, Solo Cup Company, Urbana, IL) along with one P. maculiventris (second instar, third instar, fourth instar, fifth instar, or adult). After 24 hr, the weight of the T. molitor pupa was measured again to determine the amount of weight loss due to P. maculiventris feeding. All P. maculiventris used in this study were starved for 24 hr, allowing access to water only. This experiment was repeated 30 times for each P. maculiventris instar/stage (i.e. four different instars and two different sexes of adults) and no insect was used more than one time. Because weight loss of T. molitor by feeding of younger P. maculiventris was too small to measure, we combined and measured the weight loss of three T. molitor pupae together. A total of 30 T. molitor were used as a control (i.e. without P. maculiventris in a cup). Differences in weight loss of T. molitor pupae due to P. maculiventris feeding compared to the control were analyzed with paired t-test [22].To compare feeding potential of different stages of P. maculiventris, we adopted the concept of injury equivalency proposed by Hutchins et al . [23]. The term injury equivalency was used originally to quantify the overall amount of plant injury caused by multiple pest species with different feeding potential. Feeding equivalency can be presented as feeding equivalency coefficient that measures relative feeding potential by setting the amount of feeding by a certain insect or a stage of insect as 1.0. In our study, feeding equivalency coefficient was calculated by setting the amount of feeding by the second instar as 1.0. Data on weight loss of T. molitor was analyzed with ANOVA and difference in means due to feeding for second-fifth instars and adults (male and female) were compared with Tukey’s HSD test at 5% error rate [22].
Experiment 2: Prey acceptance and feeding effectiveness of P. maculiventris on different types of prey
An experiment was conducted to determine prey acceptance and feeding effectiveness of P. maculiventris on different types of prey. A total of six different prey types in six different families and four different orders were used in this study. The prey were fully grown larvae or nymphs including the cabbage worm Pieris rapae (Lepidoptera: Pieridae) (1.9-2.4 cm in length), the cross-striped cabbage worm Evergestis rimosalis (Lepidoptera: Crambidae) (1.7-2.4 cm in length), the Colorado potato beetle Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) (0.8-1.2 cm in length), the fall web worm Hyphantria cunea (Lepidoptera: Arctiidae) (1.9-2.5 cm in length), the Mexican bean beetle Epilachna varivestis (Coleoptera: Coccinellidae) (0.6-0.8 cm in length), and the brown marmorated stink bug Halyomorpha halys (Hemiptera: Pentatomidae) (1.0-1.2 cm in length). The prey were collected from cabbage (Brassica oleracea), kale (Brassica oleracea), potato (Solanum tuberosum), black cherry (Prunus serotina), soybean (Glycine max), and corn (Zea mays). The prey was maintained in the laboratory on leaves of their host plants until the experiment was started.
One species of prey was placed in a Petri dish (5.5-cm diameter) (Lab-Tech Plastic Petri Dish, Miles Laboratories Inc., Naperville, IL) along with one P. maculiventris (third instar, fifth instar, or adult). To collect data more accurately and precisely, the feeding activity of P. maculiventris in the arena was videotaped for 24 hr using infrared cameras (The Hawk Eye Nature Cam, West Linn, OR) connected to a 4-channel DVR (Falco Model LX-4PRO, Falco Pro Series, Taiwan). This experiment was repeated 12 times for each stage/instar of P. maculiventris with each prey species. All insects were used only once in this study. From the video footage, two behaviors were quantified to determine prey acceptance and effectiveness of P. maculiventris: numbers of each prey killed by P. maculiventris feeding and the number of attacks required prior to successful feeding by P. maculiventris for each prey. Data were analyzed with ANOVA and effectiveness of P. maculiventris killing six different prey species were compared with Tukey’s HSD test at 0.05% error rate [22].
Experiment 3: Feeding preference of P. maculiventris on three different prey types
An experiment was carried out to determine the feeding preference of three different stages of P. maculiventris (i.e. third instar, fifth instar, and adult) on three different types of larvae. We chose H. cunea, E. rimosalis, and E. varivestis as prey in this study because they were accepted by P. maculiventris as prey (see Results of Experiment 2) and they exhibited different types of body surface; H. cunea has a hairy body surface, E. rimosalis has a smooth body, and E. varivestis has a spiny body surface. One H. cunea (1.8-2.5 cm in length), one E. rimosalis (1.9-2.4 cm in length) and one E. varivestis (0.6-1.1 cm in length), was randomly assigned to one quadrant of a Petri dish (9-cm diameter) (Pyrex Petri Dishes, Science Kit and Boreal Laboratories, Tonawanda, NY). Then, one P. maculiventris was introduced into the remaining quadrant of the Petri dish. The experiment was videotaped for 24 hr using infrared cameras and a 4-channel DVR. The experiment was repeated 20-22 times for each P. maculiventris. All P. maculiventris used in this study were starved for 24 hr before the experiment and all insects were used only once. From the video footage, two different data were collected: prey that P. maculiventris initially attacked and prey that was fed upon successfully. Attacking behavior was indicated by P. maculiventris protruding its proboscis toward prey and initiating feeding, but not killing the prey. Successful feeding behavior was indicated by P. maculiventris feeding upon and killing prey after attacking. Data were analyzed using chi-square test [22] to determine the feeding preference of P. maculiventris based on the types of prey for initial attacking and successful feeding.
Results
Experiment 1: Feeding potential of different P. maculiventris stagesThe weight loss of T. molitor by P. maculiventris feeding was significantly higher than that of the control (t = 6.74-11.80; d.f. = 18; P < 0.05). Also, there was a significant difference in feeding amount among different stages of P. maculiventris (F = 38.48; d.f. = 5, 44; P < 0.05) (Table 1). The fifth instar P. maculiventris caused the largest amount of weight loss of T. molitor pupae and the second instar fed the least amount. When feeding equivalency for the second instar of P. maculiventris was designated as 1.0., feeding equivalencies for nymphs and adults were 1.83-9.45 and 5.67-6.21, respectively (Table 1). Experiment 2: Prey acceptance and feeding effectiveness of P. maculiventris on different types of prey
Experiment 2: Prey acceptance and feeding effectiveness of P. maculiventris on different types of prey
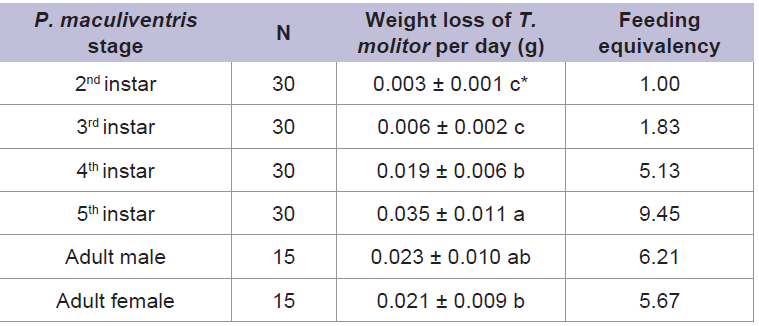
Table 1: Amount of weight loss (mean ± SEM) of T. molitor by P. maculiventris feeding and feeding equivalency when the feeding amount by second instar is set as 1.00. Note that first instar is not predacious.
Overall, P. maculiventris caused the highest mortality to P. rapae larvae (Table 2), and the lowest mortality to H. halys. The number of attacks required prior to effective feeding upon different prey species differed significantly for the third instar P. maculiventris (F = 8.05; d.f. = 4, 27; P < 0.05). Third instar P. maculiventris required six attacks to effectively feed on H. cunea and E. varivestis larvae (Figure 1). However, there were no effects of prey species on the number of attacks required prior to effective feeding for fifth instar (F = 1.88; d.f. = 4, 27; P > 0.05) and adult P. maculiventris (F = 1.70; d.f. = 4, 38; P > 0.05). P. maculiventris attempted to attack H. halys continuously throughout the experimental period, but it could not complete any effective feeding on H. halys at all due to the prey’s high mobility and chemical defense.
 Experiment 3: Feeding preference of P. maculiventris on three different prey types
Experiment 3: Feeding preference of P. maculiventris on three different prey types
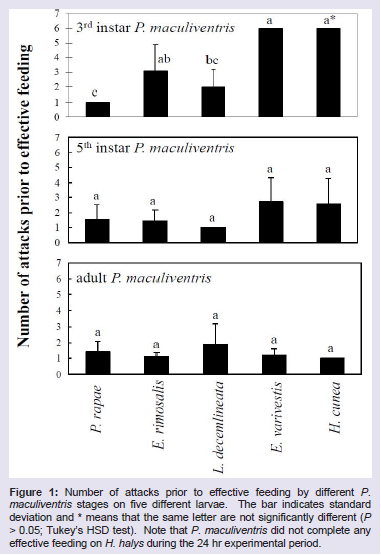
Figure 1: Number of attacks prior to effective feeding by different P. maculiventris stages on five different larvae. The bar indicates standard deviation and * means that the same letter are not significantly different (P > 0.05; Tukey’s HSD test). Note that P. maculiventris did not complete any effective feeding on H. halys during the 24 hr experimental period.
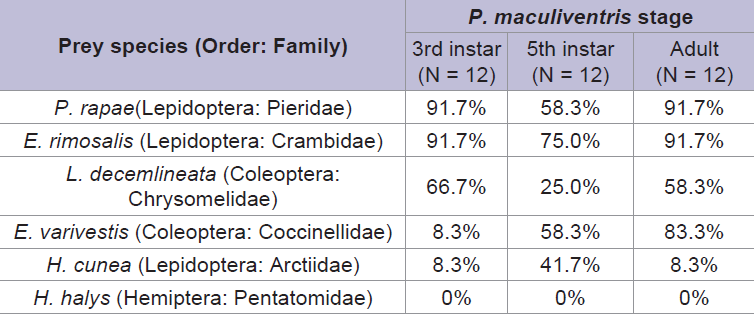
Table 2: Mortality of different prey species caused by third instar, fifth instar, and adult P. maculiventris feeding. Note that some P. maculiventris could feed and kill more than one type of larvae.
Overall, adult P. maculiventris significantly (χ2 = 13.30; d.f. = 2; P < 0.05) preferred to initially attack smooth prey (i.e. E. rimosalis) (Figure 2A). The pair-wise comparison for the first attack showed that adult P. maculiventris significantly preferred smooth prey over spiny prey (i.e. E. varivestis) (χ2 = 4.26; d.f. = 1; P < 0.05), smooth prey over hairy prey (i.e. H. cunea) (χ2 = 11.60; d.f. = 1; P < 0.05), and spiny prey over hairy prey (χ2 = 2.66; d.f. = 1; P < 0.05). Fifth instar P. maculiventris showed a significant (χ2 = 6.63; d.f. = 2; P < 0.05) preference to initially attack smooth prey. Pair-wise comparisons indicated that fifth instar P. maculiventris significantly preferred smooth prey over spiny (χ2 = 4.75; d.f. = 1; P < 0.05) or hairy prey (χ2 = 4.75; d.f. = 1; P < 0.05). However, fifth instar P. maculiventris did not show significant difference in preference between spiny and hairy prey (χ2 = 0.11; d.f. = 1; P > 0.05). Third instar P. maculiventris showed a significant (χ2 = 9.33; d.f. = 2; P < 0.05) preference to first attack smooth prey. Pair-wise comparisons revealed that third instar P. maculiventris had significant preference to initiate attack on smooth prey over spiny (χ2 = 0.81; d.f. = 1; P < 0.05) or hairy prey (χ2 = 7.00; d.f. = 1; P < 0.05). Third instar did not show significant (χ2 = 2.50; d.f. = 2; P>0.05) difference in preference to first attack between spiny and hairy prey. The results of the feeding preference test showed that adult P. maculiventris significantly (χ2 = 17.66; d.f. = 2; P < 0.05) preferred to feed upon smooth prey (Figure 2B). Pair-wise comparisons showed that adult P. maculiventris preferred smooth prey over spiny prey (χ2 = 4.48; d.f. = 1; P < 0.05), smooth prey over hairy prey (χ2 = 16.20; d.f. = 1; P < 0.05), and spiny prey over hairy prey (χ2 = 5.44; d.f. = 1; P < 0.05). Fifth instar P. maculiventris showed a significant (χ2 = 20.8; d.f. = 2; P < 0.05) feeding preference on smooth prey over hairy and spiny prey. Pair-wise comparisons revealed that fifth instar P. maculiventris significantly preferred smooth prey over spiny prey (χ2 = 7.2; d.f. = 1; P < 0.05), smooth prey over hairy prey (χ2 = 16.00; d.f. = 1; P < 0.05), and spiny prey over hairy prey (χ2 = 4.00; d.f. = 1; P < 0.05). Third instar P. maculiventris had a significant (χ2 = 6.72; d.f. = 2; P < 0.05) preference for feeding on smooth prey. Pair-wise comparisons indicated that third instar P. maculiventris showed feeding preferences for smooth prey over hairy prey (χ2 = 7.00; d.f. = 1; P < 0.05) and spiny prey over hairy prey (χ2 = 4.00; d.f. = 1; P < 0.05). However, no significance was found between smooth and spiny prey (χ2 = 0.82; d.f. = 1; P < 0.05) for third instar P. maculiventris.
The results of the feeding preference test showed that adult P. maculiventris significantly (χ2 = 17.66; d.f. = 2; P < 0.05) preferred to feed upon smooth prey (Figure 2B). Pair-wise comparisons showed that adult P. maculiventris preferred smooth prey over spiny prey (χ2 = 4.48; d.f. = 1; P < 0.05), smooth prey over hairy prey (χ2 = 16.20; d.f. = 1; P < 0.05), and spiny prey over hairy prey (χ2 = 5.44; d.f. = 1; P < 0.05). Fifth instar P. maculiventris showed a significant (χ2 = 20.8; d.f. = 2; P < 0.05) feeding preference on smooth prey over hairy and spiny prey. Pair-wise comparisons revealed that fifth instar P. maculiventris significantly preferred smooth prey over spiny prey (χ2 = 7.2; d.f. = 1; P < 0.05), smooth prey over hairy prey (χ2 = 16.00; d.f. = 1; P < 0.05), and spiny prey over hairy prey (χ2 = 4.00; d.f. = 1; P < 0.05). Third instar P. maculiventris had a significant (χ2 = 6.72; d.f. = 2; P < 0.05) preference for feeding on smooth prey. Pair-wise comparisons indicated that third instar P. maculiventris showed feeding preferences for smooth prey over hairy prey (χ2 = 7.00; d.f. = 1; P < 0.05) and spiny prey over hairy prey (χ2 = 4.00; d.f. = 1; P < 0.05). However, no significance was found between smooth and spiny prey (χ2 = 0.82; d.f. = 1; P < 0.05) for third instar P. maculiventris.
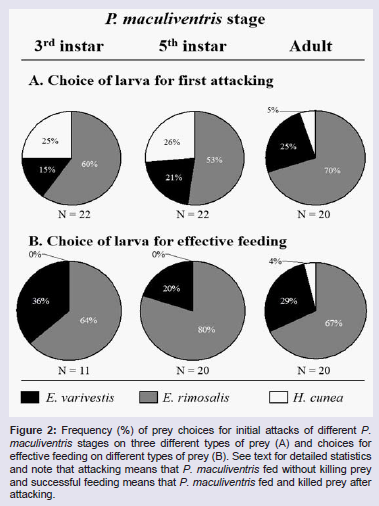
Figure 2: Frequency (%) of prey choices for initial attacks of different P. maculiventris stages on three different types of prey (A) and choices for effective feeding on different types of prey (B). See text for detailed statistics and note that attacking means that P. maculiventris fed without killing prey and successful feeding means that P. maculiventris fed and killed prey after attacking.
Discussion
Providing the optimum number of prey needed for rearing natural enemies is important because cannibalism among natural enemies can occur when less prey is offered [23] and it is economically impractical when excessive prey are offered. Wiedenmann and O’Neil [24] showed that the survival and fecundity of P. maculiventris was highly reduced if less food was offered, indicating that provision of optimal amount of prey could reduce the cost of mass rearing. In addition, adequate prey quantity is essential for optimal fitness and maximum reproductive potential for natural enemies [25]. The size of P. maculiventris increases with their age: 1.60, 2.95, 4.3, 5.8, 8.6, 10.0 mm in length for the first, second, third, fourth, fifth instars and adults, respectively [26]. Feeding amount also increases as P. maculiventris weight increases [27]. This study showed a trend that the amount of feeding by P. maculiventris increased with age except in the fifth instars. We attribute the higher amount of feeding by the fifth instars to the physiological requirement that the fifth instars need more energy for imminent morphological and physiological development of reproductive organs. Strohmeyer et al. [28] found that P. maculiventris females fed more than males, which is contradictory to our study. DeClercq and Degheele [29] also reported that mated P. maculiventris females consumed more prey than males. We hypothesize that this contradiction could be due to the mating status of the P. maculiventris females. P. maculiventris adults used in this study were virgin. Additional research would be needed to determine the effect of mating status on feeding amount of male and female P. maculiventris.The results of this study suggest two important considerations for using P. maculiventris as a generalist predator in biological control. First, the effect of P. maculiventris stage on feeding amount needs to be considered in mass rearing of P. maculiventris. This study showed a larger amount of feeding by fifth instar and adult P. maculiventris, indicating that age and size of P. maculiventris is an important factor for determining the amount of prey needed in mass rearing systems. Feeding amount of one fifth instar P. maculiventris is equivalent to ten second instars or five third instars. Because the feeding potential of different life stages of P. maculiventris could vary significantly, the feeding equivalency will also help to decide the amount of T. molitor that each stage of P. maculiventris need each day. The second, third, fourth and fifth instars of P. maculiventris last on average for 5.5, 5, 5 and 9.5 days, respectively [20]. Our study indicates that the mass rearing of one P. maculiventris from second through fifth instar will require ca. 14 T. molitor. Second, although P. maculiventris is known to prefer to feed on larvae with lower behavioral defenses [19], this study showed that morphological characteristics of prey also can deter or hinder P. maculiventris feeding. Although our study was not designed to investigate chemical defense of prey against P. maculiventris in detail, we observed that H. halys could avoid effective feeding by P. maculiventris not only because it was highly mobile but because it used chemical defenses when P. maculiventris tried attacking.
Acknowledgments
We thank V. Kondo, M. McKinney, V. Varghese, and S. Baek for their help with collecting experimental insects and rearing P. maculiventris for this study. This research was supported by USDA Organic Research and Education Initiatives Grant, and Cooperative Research Agreement with Rural Development Administration of Korea.References
- Bale JS, van Lenteren JC, Bigler F (2008) Biological control and sustainable food production. Philos Trans R Soc Lond B Biol Sci 363: 761-776.
- Coppel HC, Mertins JW (1977) Biological insect pest suppression. Springer- Verlag, Berlin, Germany.
- Symondson WOC, Sunderland KD, Greenstone MH (2002) Can generalist predators be effective biocontrol agents? Annu Rev Entomol 47: 561-594.
- Edwards CA, Sunderaland KD, Gerorge KS (1979) Studies on polyphagous predators of cereal aphids. J Appl Ecol 16: 811-823.
- Chiverton PA (1986) Predator density manipulation and its effect on populations of Rhopalosiphumpadi (Hom.: Aphididae) in spring barley. Ann Appl Biol 109: 49-60.
- Lövei GL, Sunderland KD (1996) Ecology and behavior of ground beetles (Coleoptera: Carabidae). Annu Rev Entomol 41: 231-256.
- Cottrell TE, Yeargan KV (1998) Effect of pollen on Coleomegilla maculata (Coleoptera: Coccinellidae) population density, predation, and cannibalism in sweet corn. Environ Entomol 27: 1402-1410.
- Jacometti M, Jorgensen N, Wratten S (2010) Enhancing biological control by an omnivorous lacewing: floral resources reduce aphid numbers at low aphid densities. Biol Contr 55: 159-165.
- Stoner A, Metcalfe AM, Weeks RE (1975) Plant feeding by Reduviidae, a predacious family (Hemiptera). J Kansas Entomol Soc 48: 185-188.
- Wyss E (1995) The effects of weed strip on aphids and aphidophagous predators in an apple orchard. Entomol Exp Appl 75: 43-49.
- Koch RL (2003) The multicolored Asian lady beetle, Harmonia axyridis: a review of its biology, uses in biological control, and non target impacts. J Insect Sci 3: 1-16.
- Eubanks MD, Denno RF (2000) Health food versus fast food: the effects of prey quality and mobility on prey selection by a generalist predator and indirect interactions among prey species. Ecol Entomol 25: 140-146.
- Richards LJ (1982) Prey selection by an intertidal beetle: field test of an optimal diet model. Oecologia 55: 325-332.
- Dyer LA, Floyd T (1993) Determinants of predation on phytophagous insects: the importance of diet breadth. Oecologia 96: 575-582.
- Cogni R, Freitas AVL, Amaral Filho F (2002) Influence of prey size on predation success by Zeluslongipes L. (Het., Reduviidae). J Appl Entomol 126: 74-78.
- Foglar H, Malausa JC, Wajnberg E (1990) The functional response and preference of Macrolophus caliginosus (Heteroptera: Miridae) for two of its prey: Myzus persicae and Tetranychus urticae. Entomophaga 35: 465-474.
- McPherson JE (1980) A list of the prey species of Podisus maculiventris (Hemiptera: Pentatomidae). Great Lakes Entomol 13: 18-24.
- Waddill V, Shephard M (1975) Dispersal of Podisus maculiventris nymphs in soybeans. Environ Entomol 4: 233-234.
- Marston NL, Schmidt GT, Biever KD, Dickerson WA (1978) Reaction of five species of soybean caterpillars to attack by the predator Podisus maculiventris. Environ Entomol 7: 53-56.
- Mukerji MK, LeRoux EJ (1965) Laboratory rearing of a Quebec strain of the pentatomid predator, Podisus maculiventris (Say) (Hemiptera: Pentatomidae). Phytoprotection 46: 40-60.
- Hough-Goldstein JA, McPherson D (1996) Comparison of Perillus bioculatus and Podisus maculiventris (Hemiptera: Pentatomidae) as potential control agents of the Colorado potato beetle (Coleoptera: Chrysomelidae). J Econ Entomol 89:1116-1123.
- SAS Institute (2008) SAS OnlineDoc® version 9.1.3. SAS Institute. Cary, NC.
- Hutchins SH, Higley LG, Pedigo LP (1988) Injury equivalency as a basis for developing multiple-species economic injury levels. J Econ Entomol 81: 1-8.
- Pires EM, Zanuncio JC, Serrão JE (2011) Cannibalism of Brontocoristabidus and Podisusnigrispinus during periods of prerelease without food or fed with Eucalyptus cloeziana plants. Phytoparasitica 39: 27-34.
- Wiedenmann RN, O’Neil RJ (1990) Effects of low rates of predation on selected life-history characteristics of Podisus maculiventris (Say) (Heteroptera: Pentatomidae). Can Entomol 122: 271-283.
- Omkar, Sahu J, Kumar G (2010) Effect of prey quantity on reproductive and developmental attributes of a ladybird beetle, Anegleis cardoni. Int J Trop Insect Sci 30: 48-56.
- McPherson JE (1982) The Pentatomoidea (Hemiptera) of northeastern North America with emphasis on the fauna of Illinois. Southern Illinois University Press, Carbondale, IL.
- Strohmeyer HH, Stamp NE, Jaromski CM, Bowers MD (1998) Prey species and prey diet affect growth of invertebrate predators. Ecol Entomol 23: 68-79.
- Gyawaly S (2011) Feeding behavior of Podisus maculiventris (Say) (Hemiptera: Pentatomidae) implications for mass rearing and biological control. West Virginia University, Morgantown, WV.
- De Clercq P, Degheele D (1997) Effects of mating status on body weight, oviposition, egg load, and predation in the predatory stinkbug Podisus maculiventris (Heteroptera: Pentatomidae). Ann Entomol Soc Am 90: 120-127.
- Dyer LA (1997) Effectiveness of caterpillar defenses against three species of invertebrate predators. J Res Lepid 34: 48-68.