
Journal of Plant Biology & Soil Health
Download PDF

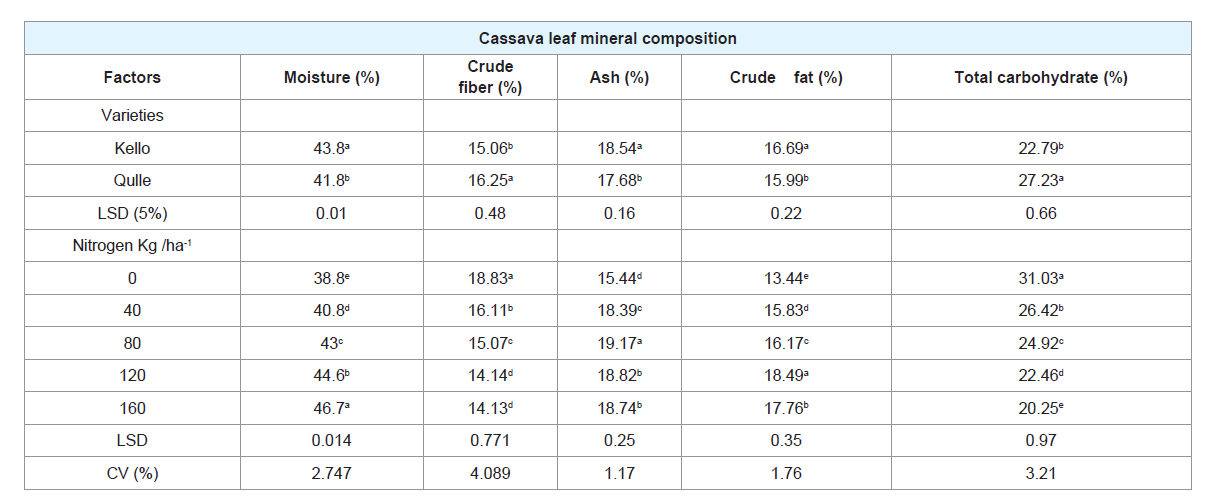 Table 1: Effect of varieties and rates of nitrogen on proximate leaf composition of two cassava varieties.
Table 1: Effect of varieties and rates of nitrogen on proximate leaf composition of two cassava varieties.
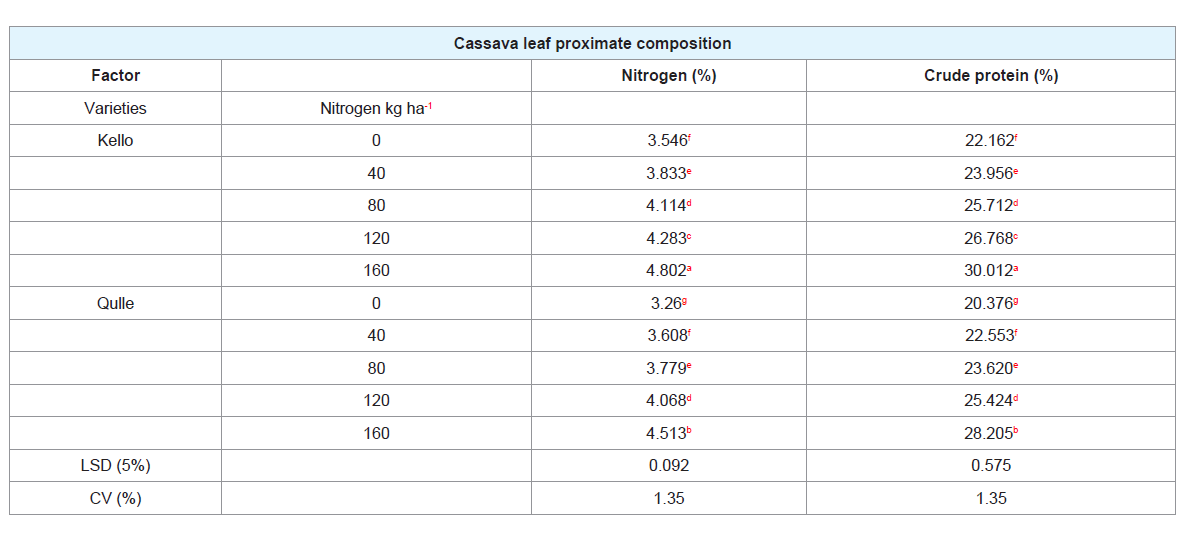 Table 2: Interaction effect of varieties and nitrogen rates on nitrogen and crude protein content of two varieties of cassava leaf.
Table 2: Interaction effect of varieties and nitrogen rates on nitrogen and crude protein content of two varieties of cassava leaf.


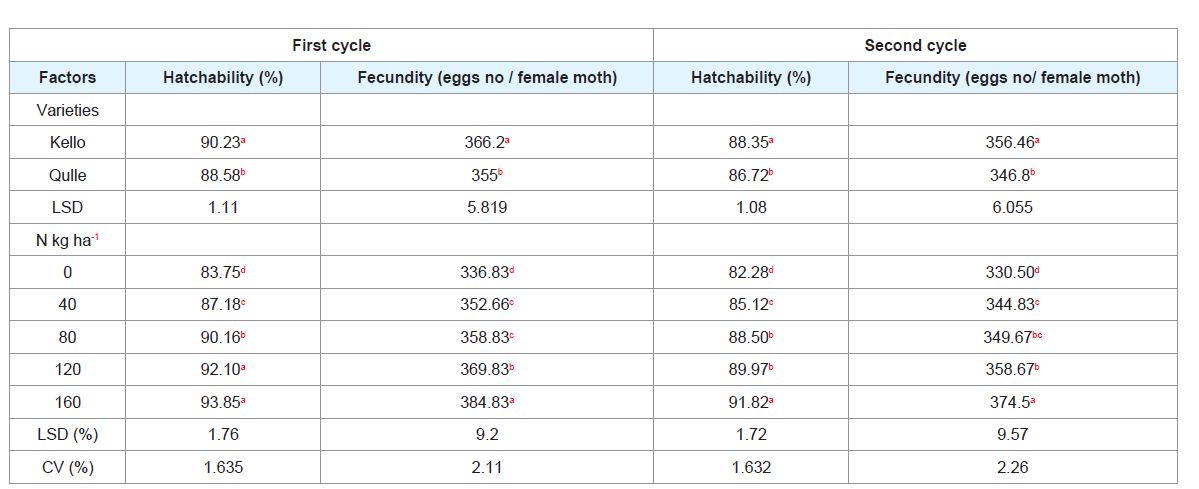 Table 3: Effect of varieties and nitrogen rates on grainage parameters of eri-silkworm.
Table 3: Effect of varieties and nitrogen rates on grainage parameters of eri-silkworm.
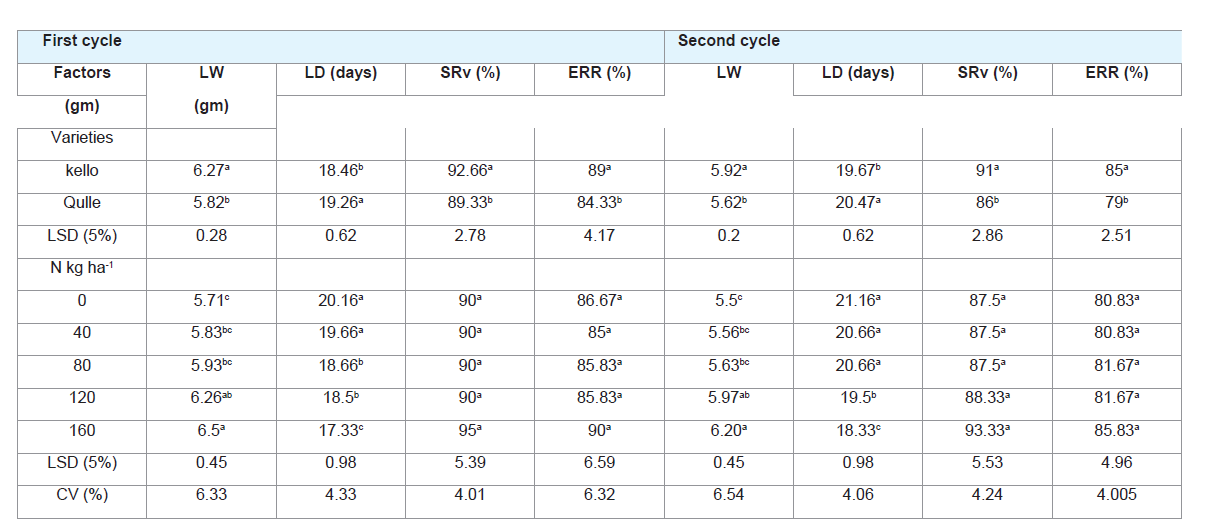 Table 4: Effect of varieties and nitrogen rates on larval parameters of eri-silkworm.
Table 4: Effect of varieties and nitrogen rates on larval parameters of eri-silkworm.

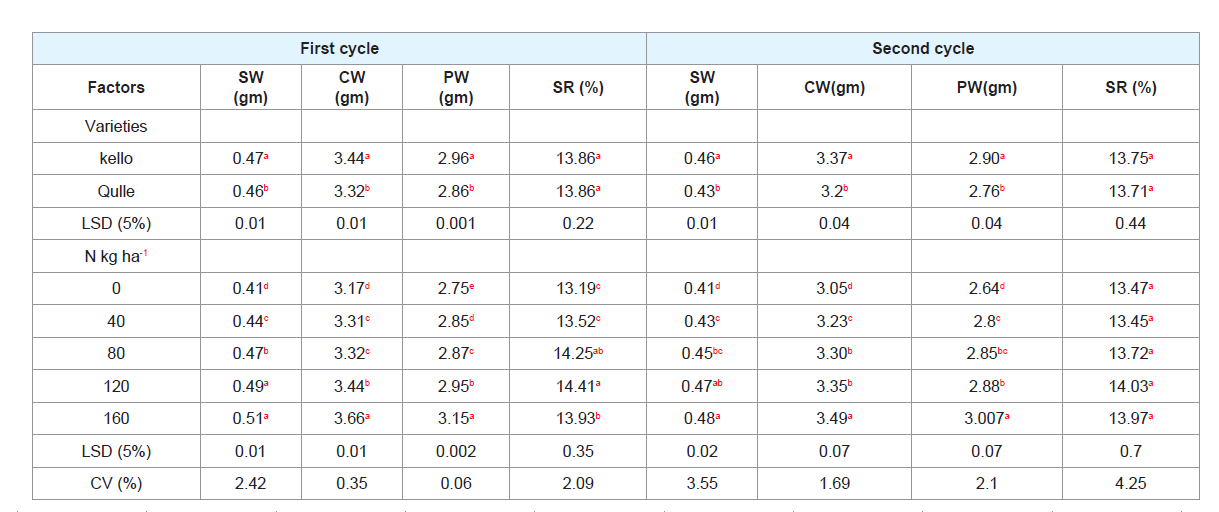 Table 5: Effects of varieties and nitrogen rates on cocoon parameters of eri-silkworm.
Table 5: Effects of varieties and nitrogen rates on cocoon parameters of eri-silkworm.
Research Article
Effect of Nitrogen Rates on Leaf Quality of Two Cassava (Manihot Esculenta Crantz) Varieties as a Feed for Eri-Silkworm (Samia Cynthia Ricini Boisduval) Cocoon Production at Jimma, Southwest Ethiopia
Derara Y1*, Sori W2, Nebiyu A3 and Mulat F3
1Department of Plant Sciences, Salale University, Ethiopia
1Department of Plant Sciences, Jimma University, Ethiopia
1Department of Plant Sciences, Mekdela University, Ethiopia
*Address for Correspondence: Derara Y, 1Salale University, Department of Plant Sciences, P.O box 245, Salale, Ethiopia; E-mail: johnderara4@gmail.com
Submission: November-26-2019
Accepted: January-02-2020
Published: January-04-2020
Copyright: © 2020 Derara Y, et al. This is an open access article
distributed under the Creative Commons Attribution License, which
permits unrestricted use, distribution, and reproduction in any medium,
provided the original work is properly cited.
Abstract
Limited research efforts have been made on cassava leaf usage for rearing eri-silkworm in relation to nitrogen fertilizer management. Hence, field and laboratory experiments were conducted at Jimma
University (Ela-dale Site) during 2016 with the objective of determining the effect of nitrogen on leaf quality and identifying appropriate cassava variety for better leaf quality as a feed for Eri-silkworm performance. A 2x5 factorial experiment arranged in Randomized Complete Block Design (RCBD) with three replications was used. The treatments consisted of two cassava varieties (Kello and Qulle) and five levels of N (0,40,80,120 and 160 kg N ha-1) as Urea. For the laboratory experiment, to rear eri-silkworm, cassava leaves harvested from the field experiment (10 plots) were arranged in Completely Randomized
Design (CRD) with three replications and their effect on Eri-silkworm performance assessed in terms of larval, cocoon, and post cocoon traits. Data were collected on growth attributes, leaf proximate
analysis of cassava, eri-silkworms larval, and cocoon and post cocoon traits. Variety and nitrogen had variable in both laboratory and fields experiment including biochemical composition of leaves. Variety Kello
was found to be superior compared to Qulle by revealing a silkworm rearing performance indicators of hatchability (90.23%), fecundity (366.2 eggs/female), shorter larval duration (18.46 days), better larval
weight (6.27 gm), survival rate (92.66%), cocoon weight (3.44 gm), single shell weight (0.47 gm), pupal weight (2.96 gm) and effective rate of rearing (89%). Application of 160 kg N ha-1 gave highest hatchability (93.85%), fecundity (384.83 eggs per female), larval weight (6.5 gm), shell weight (0.51 gm), cocoon weight (3.66 gm), pupal weight (3.15 gm) and shortest larval duration (17.333 days). In terms of biochemical composition, kello leaves had higher moisture content (0.438%), ash (18.541%) and crude fat (16.69%). Higher crude fiber (16.248%) and total carbohydrate (27.234%) were obtained from Qulle variety. 160 kg N ha-1 gave the highest moisture (0.467%) and the lowest crude fiber (14.123% ). The highest leaf nitrogen content (4.802%) and crude protein (30.012%) was gained from the combination of kello and 160 kg N ha-1. In conclusion, variety Kello and 160 kg N ha-1 can be used for higher leaf yield, leaf quality and cocoon yield.
Keywords
Nitrogen rates; Cassava leaf; Samia cynthia ricini; Biochemical
composition; Cocoon traits
Introduction
In Ethiopia, agricultural production is of a subsistence nature.
The agricultural production system is mainly rain fed and traditional,
which is characterized by low input of improved seeds, fertilizer,
pesticides and other technologies [1]. Poverty and increasing population pressure led to decline in land holding per household
that eventually resulted in low level of production to meet even the
consumption requirement of the households [2]. Poverty reduction
and Increases in the unemployment rate due to increases in the
population, therefore, requires income generation activities like mass
rearing of silk producing organisms to obtain silk [3].
More than 85% of the Ethiopian population, which resides in the
rural area, is engaged in agricultural production as a major means
of livelihood [4]. The Ethiopian government’s development policy
emphasizes agricultural sector development led industrialization. In
1996, the government initiated a food security strategy built around,
increasing agricultural production at the household level, ensuring
access to food for food deficit households, and strengthening
institutional emergency response capabilities to achieve food security
[5,6].
Sericulture provides gainful employment, economic development
and improvement in the quality of life to the people in rural area and
therefore it plays an important role to meet this policy. It provides
employment at various levels [7]. In his review of sericulture
Industry in India classified the employment generation pattern of the
industry into two major types: (i) Direct Employment - (a) Mulberry
Cultivation; (b) Leaf Harvesting; (c) Silk Worm Rearing; (ii) Indirect
Employment - (a) Reeling; (b) Twisting; (c ) Weaving; (d) Printing &
Dyeing; (e) Finishing; (f) Silk Waste Processing [8];
Sericulture is an agro-based industry. Sericulture provides economic development and improvement in the quality of life to the people in rural area and therefore it plays an important role in anti-poverty programme and prevents migration of rural people to urban area in search of employment. It can make employment up
to 11 persons for every kilogram of raw silk produced; out of which more than 6 persons are women [9]. Estimated that one hectare of mulberry creates employment of 13-16 persons per year and their location specific analysis indicated that for a production of one kilogram of raw silk, 11 man days are required which can, in turn, employ 30 man days for production of silk fabric. Showed that 96.36 man days of employment are generated from the establishment of one acre of mulberry garden for rearing 300 dfls (disease free layings) of silkworms in two months [10].
Historical evidence show that silk was discovered in China and
from there it spread to other parts of the world. Silk is produced
by the larvae from a variety of insects and spiders of the phylum
Arthropoeda. It is a natural protein fiber and is very soft, lustrous,
smooth, strong and durable than any natural or artificial fiber [11].
Silkworm is a kind of insect which can produce silk solution. Insects
mainly belong to two families, viz., Saturnidae and Bombycidae,
which spins silk fibre [12]. Among saturniidae family S.c ricini is
the one commercially exploited silkworm species and can be reared
in doors throughout the year to produce silk [13,14]. Bombyx mori
belongs to Bombycidae produces a delicate twin thread of silk fibroin,
which is coated by a protective cover of sericin. Silk protein is a kind
of protein like collagen, elastin, keratin, fibroin, sporgin etc., is an
essential constituent of cocoon filament [15]. Reported that the silk
fiber is almost a pure protein fiber composed of two types of proteins
viz., sericin and fibroin [16].
Cassava, the most preferred food plant after castor has also been
proved to be suitable for commercial rearing of silkworm. Cassava
growers can divert a portion of foliage (25-40%) to raise eri silkworm
and get extra income without affecting the tuber yield and starch
content [17-19]. Also stated that cassava cultivation for eri silkworm
rearing provides gainful employment to the women [20].
Eri silkworm is a domesticated multivoltine non-mulberry
silkworm. It is a sericigenous insect exploited for its valuable eri silk.
It is a multivoltine insect completing at least six to seven generations
in a year and it can be reared throughout the year depending on
the availability feed [14,21]. The agro-ecology and feed availability
are the major requirements which have significant effect on rearing
of larvae of this insect and finally cocoon crop yield and quality.
It is a domesticated silkworm that feeds primarily on leaves of the
castor plant. Eri-silkworm shows different rearing behavior when
fed different food plants [22]. Eri-silkworm rearing also relies on
management practices i.e., rearing temperature, humidity and
nutrition. The healthier rearing conditions, environment and
nutrition during larval period may lead to higher fecundity by
silkworm moths [23,24].
The quality of feed plays a very important role for growth and
development of the eri-silkworm and ultimately on the economic traits
of cocoons. Leaf is a key factor besides environment and technology
adoption for better growth and development of the silkworm larvae
and cocoon production [25-27]. Reported that, among the various
factors influencing silkworm growth and cocoon production, leaf
quality plays a major role [28]. Eri- silkworm fed with more moisture,
protein, sugar and carbohydrates and less minerals and crude fibre
content is the best from the silkworm nutrition point of view [29].
Nitrogen application influences the quality of feed (leaf) especially
its protein content [30,31]. As nitrogen is an important limiting factor
for phytophagous insects, reduction of nitrogen contents has forceful
effects on insect performances. The weights of larvae and cocoons
of eri-silkworm are significantly influenced by nitrogen content of
foliage [32]. Deficiencies in Nitrogen can also affect cassava yield in many ways. This might occur through reduction in leaf area,
moderate reduction in photosynthetic rates of leaves, modification
of branching habit and changes in the distribution patterns of dry
matter to different plant parts [33].
Therefore, nitrogen application is very important for eri silkworm
performance and cocoon production. In line with this, the present
study was carried out with the general objective of evaluating
performance of eri-silkworm reared on leaf of two cassava varieties
cultivated under different rates of nitrogen application.
Materials and Methods
Description of the study area:
The experiment was conducted at the experimental field in Jimma
University College of Agriculture and Veterinary Medicine, Eladale
Site, during 2015/2016 rainy season. It is about 365 km far from
Addis Abeba on the road to Bedele. The research site is geographically
located at 7o 42` N latitude and 36o 50`E longitude with an altitude of
1710 m above sea level. The dominant soils of the area are Nitisol and
Cambisol. The area receives an average annual rainfall of 1530 mm.
The area has an average maximum and minimum temperature of 26.2
oC and 11.3 oC, respectively and average maximum and minimum
relative humidity of 91.40% and 37.92%, respectively [34].
Agriculture is the main occupation of the area and raising
silkworm for eri cocoon production is not widely practiced but
recently people in the study area are familiarized with silkworm
production as a source of income.
Experimental materials and treatments:
Two cassava varieties, namely Kello and Qulle, which were
developed and released in 2005 by Hawassa Agricultural Research
Centre, were used for this experiment. They were chosen for the
study because of the fact that they have a wide range of adaptation
and are high yielders and popular among farmers in the south and
southwestern part of Ethiopia.
Eggs (seeds) of white plain 2.4 breed of eri-silkworm were used
to study the effect of varieties and rates of nitrogen on eri-silkworms
larval growth, cocoon and post cocoon characters. This type of erisilkworm
is hardy, disease resistant and can easily be reared [3].
The treatment consisted of factorial combinations of two
cassava varieties (Kello and Qulle) and five different nitrogen rates
(0,40,80,120 and 160 kg ha-1) in the form of urea (46% N). Each level
of nitrogen was combined with two levels of varieties. The national
blanket fertilizer recommendation for cassava (80 kg N ha-1) was used
as a bench mark in the experiment.
Experimental designs:
A. Field experiment: The field experiment to study the effect of nitrogen rates on leaf quality of two cassava varieties consisted of factorial combinations of two cassava varieties (Kello and Qulle)
and five different nitrogen rates (0, 40, 80, 120 and 160 kg ha-1) laid out in Randomized Complete Block Design forming 10 treatment combinations each replicated thrice. Gross plot size was 5m x 4m (20 m2) and the stem cuttings were planted at the spacing of 1m between rows and 1m between plants. There were five rows per a single plot and four stem cuttings were planted per row. The total number of stem cuttings per a single plot was therefore 20. The distance between plot and block was 1 meter and 1.5 meters, respectively leaving the outermost rows on both sides of each plot to avoid border effects. The entire experimental area was 882 m2.
B. Laboratory eri-silkworm rearing experiment:
A laboratory experiment consisting of leaves of two cassava varieties (Kello and
Qulle) grown under five rates of nitrogen (0,40,80,120,160 kg N ha-1)
were arranged in Completely Randomized Design (CRD) to form 10
treatment combinations each replicated thrice. Therefore, there were
30 plastic trays in the rearing room which represented the plots on
the field. A single plastic tray contained 20 worms. Total worms on
trays were 600.
Experimental procedures:
A. Field experiment: The experimental field was prepared
following the conventional tillage practice before planting the stem
cuttings. The stem cuttings of about 30 cm long were planted on
ridges at 450 according to the recommended space of 1 m by 1 m
during the onset of the main rainy season. Urea (46% N) fertilizer
was used as a source of nitrogen. The nitrogen fertilizer was divided in
two equal splits and the first half was applied during first month and
the remaining half was side dressed three month after planting [35].
B. Laboratory eri-silkworm rearing experiment: Eggs of erisilkworm
were procured from Jimma Agricultural Research Center.
Rearing room and appliances was cleaned before starting rearing eri
silkworm with 2% formalin solutionto protect pathogens which infect
eri silkworm [36,37]. The room was kept open for a minimum of 12
hours to remove the traces of formalin vapor left over in the room
before the starting of the rearing experiments. The eggs procured
were subjected to surface sterilization with 2% formalin solution and
washed with clean water and dried under shade to ensure proper
hatching [38]. After drying, the eggs were incubated at normal room
temperature till hatching. The newly hatched larvae were selected and
transferred with the help of forceps to rearing tray.
Rearing was undertaken as per the standard rearing technique
under laboratory conditions from first to fifth instar on (Jan-March)
and (April-June) [39,40]. Fresh and healthy leaves of Kello and Qulle
varieties of cassava were used in the present study. The leaves were
harvested daily from the cassava garden during the early hours of the
day and stored cool to maintain its freshness. The first and second
instar larvae were fed with tender leaves; medium aged leaves were
fed to the third instar larvae and matured leaves to the fourth and fifth
instar larvae. The larvae were continuously fed at a frequency of four
times per day at intervals of six hours except during molting periods
[42]. The quantity of food was increased with the advancement of larval
age to fulfill their requirement. Bed cleaning was done to ensure the
cleanliness in the immediate locality of silkworms in order to protect
from disease infection and to ensure them good feeding appetite
[43]. Adequate ventilation was ensured to the larvae by placing
the trays one above the others crosswise. The matured larvae were
relocated to suitable mountages separately for spinning of cocoons
[44]. Cocoon harvesting was done after the sixth day of spinning [45].
This procedure was conducted for two life cycles of silkworm due to
the fact that the previous feed of the eggs of eri-silkworm brought from Jimma agricultural research center was castor. Data on grainage
parameters (hatchability and fecundity), larval parameters (larval
duration, larval weight, survival rate and effective rate of rearing)
and cocoon parameters (shell weight, cocoon weight, pupal weight
and silk ratio) were assessed, recorded and statistical analysis was
performed using standard tools.
Data collected:
Leaf proximate analysis: The leaf samples at three different
heights of the plant viz., top, middle and bottom, were collected in
paper bags at 180 days after stem planting and composite leaf samples
were made. Leaves were shade dried for three days and then dried in
hot air oven at 70 oC until constant weight was gained. The dried leaf
samples were ground into fine powder and well-maintained in butter
paper bags for chemical analysis. Each sample had three replications.
As adopted by [46], crude protein was estimated by multiplying the
estimated value of the total nitrogen by 6.25, while the total nitrogen
content of the leaf was determined by Kjeldahl method. Techniques
and procedures of A.O.A.C. (2000) were used to determine crude
fiber and crude fat contents [47]. The method of was used to estimate
total minerals (ash) composition and total carbohydrates [48].
Accordingly, total carbohydrate was determined by the method of
subtracting the percentage values of protein, fat, ash and fiber from
100.
Eri-silkworm characteristics: Grainage Parameters: Hatchability (%)::
After complete hatching (third day from the
beginning of hatching) the laying was counted to collect the data in
respect to the total number of eggs laid per female moth, number of
unfertilized eggs and number of hatched eggs per layings. The average
hatching of five layings was taken as representative hatchability
percentage per layings in this case [45].
B. Fecundity (eggs/ female moth):
Pairs of freshly emerged moth
were placed on mountage and kept hanging on a wire. There were
three replications of five pairs of moths in each replication for each
treatment tested. The moths were decoupled after three hours and
the female moths were allowed to lay eggs on the mountage. After
three days, the eggs laid were separated from the mountage and
counted replication wise to find out the fecundity and was recorded
as numbers of egg per female.
Larval parameters:
A. Larval duration (days): The larval duration is the period between hatching of eggs and maturity of the larvae and was recorded in each treatment replication wise in days.
B. Larval weight (gm):
The weight of 5th in star larvae was taken
when the larvae stopped eating; body became pale and excreted the
last excreta. Five larvae were randomly picked from each treatment
and weighed treatment and replication wise and their average was
calculated.
C. Survival rate of larvae (%):
This was measured by dividing
the number of survived larvae by number of larvae brushed and
expressed in terms of percent [45].
D. Effective rate of rearing (%):
This is the number of larvae spinning cocoons out of the number of larvae brushed and is expressed in percentage [45].
Cocoon parameters:
A. Single cocoon weight (gm): On the sixth days of spinning, five
cocoons were randomly harvested from each treatment and individual cocoon weight was weighted and their average was recorded [45].
B. Shell weight (gm):
Randomly selected five cocoons were cut
open, their pupae and larval excuvium were removed and average
shell weight was recorded separately.
C. Pupal weight (gm):
Similarly, after the cocoons were cut open, single pupa weight was recorded using sensitive balance. It can also be obtained by subtracting the shell and exuviae weight from the cocoon weight.
D. Shell ratio (%):
The amount of silk present in a cocoon shell was expressed in percentage. It is weight of cocoon shell out of weight of the cocoon with pupa [39].
Data Analysis:
All data collected on different parameters of proximate compositions of leaf, grain age, larval and cocoon parameters were first checked for normality and did not violate the rule. The data were subjected to analysis of variance (ANOVA) using SAS version 9.2 [49]. Significance differences between treatment means were delineated by LSD (Least Significance Difference) test at 5% level of significance
Results and Discussion
Effect of Varieties and Nitrogen Rates and their Interaction on Leaf Proximate Composition of Cassava Varieties:
A. Leaf moisture (%): Leaf moisture percentage of the two
cassava varieties varied highly significantly with higher content
being from Kello (43.8%) as compared to qulle (41.8%). Variation
in moisture content between varieties could be attributable for
their inherent characters. The increase in leaf moisture content
might be enhancement in hydrogen ion concentration in plant sap
due to accumulation of chlorides and less moisture loss by evapotranspiration
in the leaves [50]. These results are in conformity with
the observations of [51], Sannappa and who observed variations in
moisture content of leaves among castor genotypes [52].
From the result, application of different rates of N resulted in to
highly significantly different (P<0.001) moisture content of the leaf per plant. Moisture content of the leaf increased with the increase
in the rates of N. The leaf with highest moisture content was found
at the rate of 160 kg N ha-1 while least leaf moisture content was
documented with no application of Nitrogen (0 kg N ha-1).
B. Crude fiber (%):
The increase in N rates caused the decrease of
crude fiber from 18.829% to 14.123%. The highest crude fiber of the
leaf was documented at no (0 kg/ ha-1) nitrogen fertilizer rate while
the smallest crude fiber was observed at the highest (160 kgN ha-1)
nitrogen fertilizer treatment. This shows that increasing N fertilizer
rates decreased crude fiber in the leaf. The decline in crude fiber
content with increased N rates could probably be attributed to the
fact that plants tend to use more of their photosynthates on protein
rather than carbohydrate synthesis. In case of varieties, there was
notable variation between them. Qulle variety revealed higher crude
fiber compared to Kello variety. The variation in crude fiber between
the varieties may have occurred due to their inherent characters.
This finding is supported by the finding of who recorded significant
variation in crude fiber among castor genotypes [53]. Similar to this
experiment, also observed variation in crude fiber in different castor
genotypes [32].
C. Ash (%):
In case of ash content, higher ash content was
estimated from the leaves of kello (18.541%) variety. The main
reason for variation in ash content between the varieties could be
due to genetic factors (Table 1. Observed significant variation in ash
content between eight different castor genotypes [53]. Ash content
may also vary due to variation in metal composition in the same plant
of different variety [54]. On the other hand, application of different
rates of N highly significantly (P<0.001) influenced ash content of the
leaf per plant. Ash content of the leaf increased gradually with the
increase in the rates of N. The leaf with highest ash content (19.17%)
were recorded at the rates of 80 kg N ha-1 while least leaf ash content
(15.435%) was recorded with no N application (0 kg N ha-1). This low
ash content is indicative of the low mineral content of cassava [55].
D. Crude fat (%):
The increase in nitrogen caused an increase in crude fat of leaf. The highest crude fat in the cassava leaf (18.496%)
was registered at the rates of 120 kg N ha-1 application. Crude fat
which includes all lipids, chlorophyll, carotenes and all other fat
soluble material tends to be enhanced by N application [56]. The least
crude fat content (13.437%) was recorded with no application of N
(0 kg ha-1). Crude fat content between two cassava varieties varied
highly significantly with higher content being from kello as compared
to qulle (Table 1). Variation in crude fat content between varieties
could be attributable for their difference in terms of their inherent
characters. Consistent with the current findings, findings of observed
variation in fat content among castor genotypes [32,57].
E. Total carbohydrate (%):
The total carbohydrate which is
analyzed on dry weight basis as a difference of the sum of ash, crude
protein, crude fat and crude fiber from 100 also revealed significant
variation for different rates of nitrogen and two cassava varieties.
Qulle recorded higher carbohydrate content of 27.234% while Kello
registered 22.797%. The difference in total carbohydrate content
between the two varieties might be attributed to the variation in
genetic factors. The present observation is in agreement with the
findings of and who detected variation in the total carbohydrate
content among castor genotypes [58-60]. With regard to N, the
highest total carbohydrate was gained from application of 0kgN ha-1.
Limited N supply results in higher level of carbohydrates [56,61]. The lowest total carbohydrate was obtained from the highest application
of N (160 kg/ ha-1).
F. Leaf nitrogen (%):
The highest leaf nitrogen was recorded
from Kello variety with the application of 160 kg N ha-1 while the
lowest nitrogen content was obtained from qulle with no application
of N (Table 2). Leaf nitrogen content ranged between 3.403% and
4.657%. There was also variation between two cassava varieties.
Kello had higher nitrogen content in its leaf than Qulle. This result
is in agreement with the findings of [62]; who observed significant
difference among castor genotypes in terms of nitrogen content. In
the same manner [32,63], also recorded different amount of nitrogen from different varieties of castor [64].
G. Crude protein (%):
From the interaction point of view, the
highest leaf crude protein (30.012%) was recorded from kello variety
with the application of 160 kg N ha-1 while the F. Leaf nitrogen (%):
The highest leaf nitrogen was recorded from Kello variety with the
application of 160 kg N ha-1 while the lowest nitrogen content was
obtained from qulle with no application of N (Table 2). Leaf nitrogen
content ranged between 3.403% and 4.657%. There was also variation between two cassava varieties. Kello had higher nitrogen content in its leaf than Qulle. This result is in agreement with the findings of who observed significant difference among castor genotypes in terms of nitrogen content [32,62,63]. In the same manner, also recorded different amount of nitrogen from different varieties of castor [64].
G. Crude protein (%):
From the interaction point of view, the
highest leaf crude protein (30.012%) was recorded from kello variety
with the application of 160 kg N ha-1 while the lowest crude protein
(20.376%) was obtained from Qulle with no application of nitrogen
(Table 2). Higher crude protein content was higher for Kello variety.
The variation in the crude protein content between the two varieties
may at least partly be attributed to genetic factor. Recorded variation
in crude protein content among castor genotypes [51,52,58-60].
Crude protein which is 6.25 times the nitrogen content (Lord,
1968) varied markedly among different rates of nitrogen. The lowest
crude protein was gained from no application of nitrogen (0 kg N ha-1)
whereas the highest crude protein was obtained from the application
of the highest nitrogen (160 kg N ha-1).The possible reason for the
increment of protein content as the application of nitrogen increase
could be due to the fact that N is utilized to synthesize amino acids,
which in turn form proteins [30,31].
Effect of Varieties and Nitrogen Rates on the Rearing Performance of Eri-silkworm:
Grainage parameters: Significant differences were observed in grainage parameters of eri-silkworms when fed with cassava leaves of two varieties grown on different rates of nitrogen fertilizers (Table 3).
A. Hatchability (%):
During first and second rearing, the highest hatchability was observed from application of 160 kg N ha-1 which was statistically similar with application of 120 kg N ha-1 in the first cycle of rearing while the lowest was recorded from no application of nitrogen (0kgN ha-1). The probable reason for the increment of hatchability could be due to the increase of leaf nitrogen. The reduction of hatchability during second cycle might be due to the reduction of nutrients in the leaf, in most case, was matured/aged.
In case of varietal impact on eggs hatchability, the higher
hatchability was gained from kello variety while the lower was
attained form qulle variety. The possible reason for the difference
on hatchability could be due to the variation in floral nutrient
composition which has been shown above under the section
proximate composition analysis. Better nutrient composition of
the leaf results to better growth of the larvae leading to health moth
and eggs produced from such moth hatches normally. The present
result is corroborated with the observations of who found variation
in hatchability due to the castor genotypes when used for rearing erisilkworm
[65-68].
B. Fecundity (eggs/ female moth):
Both first and second cycle
of rearing indicated that higher fecundity was recorded from kello
variety when compared to qulle variety. The variation in fecundity
during first and second cycle with two cassava varieties may be due to
the differences in foliar compositions which contribute to the growth
and development of silk worms. This is confirmed by the findings
of who observed variation in fecundity between different castor
genotypes [69,70].
N application significantly influenced the fecundity of erisilkworm
moth. Fewer egg numbers were recorded from the control
treatment while the highest fecundity was obtained from the
application of 160 kg N ha-1 (Table 3). Fecundity increased as nitrogen
level increased. The reduction of fecundity during second cycle might
be due to the reduction of nutrients in the leaf as the leaf matured.
Larval parameters:
Significant differences were observed in larval parameters of erisilkworms
between the two varieties and different rates of nitrogen in
both rearing cycles except for survival rate of the larvae and effective
rate of rearing to different rates of nitrogen (Table 4).
A. Larval duration (days):
The longest larval duration was
recorded from zero application of N (0 kg/ha-1) which was not
statistically different from 40 kg/ha-1 nitrogen fertilizer rate. The
shortest larval duration was recorded at 160 kg N ha-1. This showed
that larval duration decreased with increase in N fertilizer. Observed
negative correlation of nitrogen with larval duration.
Significantly shorter total larval duration was recorded in
the larvae fed on Kello variety in rearing cycles, 18.47 and 19.67,
respectively. The possible reason for the variation observed in larval
duration between the two varieties might be due to their nutritional
difference. Better nutrient compositions in the leaves result in to
fast growth of the worms and reduce larval duration. Similar results
were obtained by who reported that different castor varieties fed to
silkworms exhibited differences in larval duration [66,68].
B. Matured larval weight (gm):
Larval weight varied very
significantly in two cassava varieties. Kello registered higher matured
larval weight than Qulle in both rearing cycles (Table 4). The variation
in matured larval weight of silkworm fed with two cassava varieties
might be due to nutrional difference of leaves which contribute to
the growth and development of silkworms. The characters of cocoon
primarily depend upon the larval weight. The result of matured larval
weight indicated that the feed had significant effect on them. Larval
weight can vary depending upon the feed plants which the larvae feed
[57,71]. These authors observed differences in larval weight when eri
worms were fed with leaves of different castor genotypes.
Different rates of N application resulted in to significant
differences in larval weight. The lowest larval weight was registered
from the control treatment while the highest larval weight was
obtained from the application of 160 N kg ha-1 which is statistically
similar with application of 120 kg N ha-1 (Table 4). The highly
nutritious and nutrient balanced food is the main factor responsible
for healthy growth and development of any insect. Larval weight
increased as nitrogen rate increased.
C. Survival rate (%):
The analysis of variance for survival rate showed that this trait was not influenced statistically by rates of N (P
<0.05) (Table 4).
Varieties showed significant variation on survival rate of erisilkworm.
Survival rate of eri-silkworm was higher for Kello when
compared to Qulle (Table 4). The study was in line with the work
of who found significant differences in survival rate of eri-silkworms
when fed on different castor varieties [65,72].
D. Effective rate of rearing (%):
Effective rate of rearing showed
significantly different when eri-silkworm fed on two cassava varieties
in both rearing cycles (Table 4). Eri-silkworm fed on Kello recorded
higher effective rate of rearing than Qulle. Difference in effective
rate of rearing was observed among castor genotypes used as feed by
earlier workers [67,68,73-75].
The nitrogen rates didn’t affect the effective rates of rearing. The
reduction of effective rate of rearing during second cycle might be due
to the reduction of nutrients in the leaf.
Cocoon parameters:
Cocoon parameters of eri-silkworm (shell weight, cocoon weight
and pupal weight) showed significant variations when larvae were
fed with two cassava varieties applied with different rates of nitrogen.
But, the two varieties during both cycles of rearing and different rates
of nitrogen during second cycle of rearing did not show significant
differences for silk ratio (Table 5).
A. Single cocoon weight (gm):
Significant differences were
observed in single cocoon weight of larvae fed with the leaves of
two cassava varieties Significant differences were observed in single
cocoon weight of larvae fed with the leaves of two cassava varieties
grown with different rates of nitrogen in both cycles of rearing (Table
5). Cocoon weight is the most important character for productivity of
silk farming [76]. During first and second cycles of rearing, application
at 160 kg N ha-1 resulted in to significantly higher cocoon weight
(3.66 gm and 3.496 gm). The lowest was obtained at no application
of Nitrogen. The weights of cocoons are significantly influenced by
nitrogen content of foliage [32].
Higher cocoon weight was recorded from Kello variety when
compared to Qulle. This result is in agreement with the finding of who
reported that cocoon weight depends on the type of hosts provided
for feeding the larvae [77]. The occurrence of higher values of single
cocoon weight on the feed plants of these two cassava varieties in the
present study might be due to their higher larval weight gain at the
end of the larval feeding period.
B. Shell weight (gm):
It is evident that varieties of cassava had
significant effect on shell weight of the silkworm during first and
second cycle of rearing. Shell weight was found to be significantly
higher in Kello as compared to Qulle. This finding is comparable
with the reports of who opined that the shell weight varied with the
type of hosts provided at the larval stage [65]. Also reported that shell
weight varied when eri-silkworms were reared on different varieties
of cassava leaves [71].
Different rates of Nitrogen application resulted in to differences
in shell weight of eri-silkworms (Table 5). The highest shell weight
was observed from application of 160 kg N ha-1 although it was not statistically different from application of 120 kg N ha-1. The lowest was
recorded from no application of nitrogen (0 kgN ha-1). The probable
reason for the increment of shell weight as the rates of N increased
could be due to the fact that N is considered as one of the nutrients
in the leaf which highly contributes for the growth of larvae and then
better shell weight. The reduction of shell weight in the second rearing
cycle might be due to the reduction in nutritional status of leaf.
C. Pupal weight (gm):
Pupal weight differed significantly
between two cassava varieties during both cycles of rearing. During
both rearing cycles, the higher pupal weight (2.96, 2.86 gm) was
registered from Kello variety when compared to Qulle (2.90, 2.76
gm). The result of pupal weight depends on result of cocoon weight
and shell weight. This result is in conformity with the findings of who
reported that pupal weight depends on the type of hosts provided for
feeding the worms [51,65,72,78-80].
The highest N application, 160 kg N ha-1, resulted into biggest
pupal weight and the smallest pupal weight was recorded from no
application of nitrogen (0 kg N ha-1) (Table 5). The highest values
of pupal weight from the highest application of N might be due to
highest larval weight gain at the end of the larval feeding period [81-85].
D. Silk ratio (%):
Silk ratio of eri-silkworm is highly significantly
(p<0.001) affected by nitrogen fertilizer rates during both rearing
cycles (Table 5). During first rearing cycle, the highest silk ratio was
obtained from 120 kg N ha-1 which was not statistically different from
the application of 80 kg N ha-1. The lowest silk ratio was recorded from
no application of nitrogen (0 kg/ha-1) which didn’t differ statistically
from 40 kg N ha-1. During second rearing cycle, the effect of fertilizer
did not result in statistically significant (P <0.05) difference for this
parameter (Table 5) [86-89].
Conclusion and Recommendation
The results of analysis of variance revealed significant differences
in biochemical composition and rearing performance of silk worms.
It was found that varieties and rates of nitrogen showed highly
significant influences on mineral and nutrient content of leaf and erisilkworm
grainage, larval and cocoon yield parameters. Variety Kello
produced higher value for plant leaf nutrient contents (moisture, ash
and crude fat), grainage, larval and cocoon parameters (hatchability,
fecundity, larval weight, survival rate, effective rate or rearing, cocoon
weight, shell weight, pupal weight and silk ratio), while Qulle variety
bettered on crude fiber, total carbohydrate and larval duration.
N application at 160 kg N ha-1 resulted to maximum value for
proximate composition of the leaf (moisture), and for eri-silkworm
parameters (hatchability, fecundity, larval weight, single cocoon
weight, shell weight, pupal weight and shell ratio). The highest
values for crude fiber and total carbohydrate were recorded from no
application of urea. Combined application of 160 kg N ha-1 and use of
Kello cassava variety showed higher mean number for nitrogen and
crude protein. The least record in all parameters was from the control
treatment except for crude fiber and carbohydrate.
In conclusion, cassava can grow well in the study area and farmers
can benefit more by using the application of 160 kg N ha-1 which
resulted in maximum leaf yield, leaf quality and cocoon productivity
and the variety kello which is more productive in terms of leaf quality
and cocoon productivity.
However, more research should also be carried out to support the
current findings in the following areas:
1. Amount and frequency of leaf defoliation should be studied.
2. Appropriate integration of eri silkworm rearing with
cassava tuber production.
3. Other nutrients and minerals in the leaf should further be
explored in that they have relationship with rearing performance of
eri-silkworm.
4. Cost benefit analysis should to be done.
5. Similar experiments are suggested to be carried out at
different seasons and different locations because repetition of the
experiment for more seasons and locations would help us to draw
sound conclusions and recommendation.
References
15.
Dandin SB, JayaswalJ, Giridhar K (2003) Handbook of Sericulture
Technologies, CSB, Bangalore, 259.
Citation
Derara Y, Sori W, Nebiyu A, Mulat F. Effect of Nitrogen Rates on Leaf Quality of Two Cassava (Manihot Esculenta Crantz) Varieties as a Feed for Eri-Silkworm (Samia Cynthia Ricini Boisduval) Cocoon Production at Jimma, Southwest Ethiopia. J Plant Biol Soil Health. 2020;6(2): 11.