
Journal of Vaccine & Immunotechnology
Download PDF

Determination of cytokine levels



Research Article
*Address for Correspondence: Sukhbir Kaur, Professor, Department of Zoology, Panjab University, Chandigarh-160014, India, Tel: +91-172-2534215; Fax: +91-172-2541409; E-mail: puzoology@yahoo.com
Citation: Thakur A, Kaur H, Kaur S. Immunization with Live Promastigote Antigen of Leishmania donovani induces Protection against Experimental Visceral Leishmaniasis Infection in BALB/c Mice. J Vaccine Immunotechnology. 2015;2(1): 6.
Copyright © 2015 Thakur A, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Vaccine & Immunotechnology | ISSN: 2377-6668 | Volume: 2, Issue: 1
Submission: 18 August 2015 | Accepted: 01 October 2015 | Published: 05 October 2015
Reviewed & Approved by: Dr. Jung-Hyun Park, Chief of the Cytokine Biology Unit, Experimental Immunology Branch, National Cancer Institute, National Institutes of Health, USA
Keywords: Visceral leishmaniasis, Immunity, Vaccines, Low dose antigen

DTH responses

Antibody responses
Immunization with Live Promastigote Antigen of Leishmania donovani induces Protection against Experimental Visceral Leishmaniasis Infection in BALB/c Mice.
Ankita Thakur, Harpreet Kaur and Sukhbir Kaur*
- Department of Zoology, Panjab University, Chandigarh, India
*Address for Correspondence: Sukhbir Kaur, Professor, Department of Zoology, Panjab University, Chandigarh-160014, India, Tel: +91-172-2534215; Fax: +91-172-2541409; E-mail: puzoology@yahoo.com
Citation: Thakur A, Kaur H, Kaur S. Immunization with Live Promastigote Antigen of Leishmania donovani induces Protection against Experimental Visceral Leishmaniasis Infection in BALB/c Mice. J Vaccine Immunotechnology. 2015;2(1): 6.
Copyright © 2015 Thakur A, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Vaccine & Immunotechnology | ISSN: 2377-6668 | Volume: 2, Issue: 1
Submission: 18 August 2015 | Accepted: 01 October 2015 | Published: 05 October 2015
Reviewed & Approved by: Dr. Jung-Hyun Park, Chief of the Cytokine Biology Unit, Experimental Immunology Branch, National Cancer Institute, National Institutes of Health, USA
Abstract
Visceral leishmaniasis (VL) also commonly known as kala-azar is a vector borne parasitic disease which is endemic in Indian subcontinent. At present treatment of this disease relies on chemotherapy and no vaccine is available for human use. Since chemotherapy has some side-effects therefore vaccination with long lasting immunity is the major hope to control leishmaniasis. It is presumed that if a vaccine candidate can activate immune system in a similar manner to the natural course of infection, it will be more successful in generating effective immune response. Also it is known from the previous studies that few numbers of parasites are required to be present in host system for the stimulation of appropriate immune response. Based on this approach we performed our present study in which BALB/c mice were immunized with viable low (103) dose of Leishmania donovani promastigotes. Challenge infection (107) was given to all animals after three weeks of last immunization. Mice were euthanized 15 days after immunization and on 90 post infection/challenge day. A remarkable decline in parasite load was noticed in immunized animals as compared to the infected controls after challenge infection as illustrated by the reduction in parasite burden (72%). Immunized animals showed enhanced DTH responses, elevated IgG2a antibody levels and up-regulated levels of IFN-γ and IL-12 over the infected controls which imply the generation of protective immunity. The present study indicates that such whole parasite vaccines are intrinsically safer and holds a promising potential as anti-leishmanial vaccine candidate.Keywords: Visceral leishmaniasis, Immunity, Vaccines, Low dose antigen
Introduction
Visceral leishmaniasis is a neglected tropical disease identified by the World Health Organization. Around 200 000 to 400 000 new cases of VL are reported every year [1]. The threat of this devastating disease continues because of the unavailability of promising drug therapy or vaccine. Extensive research is going on to develop a promising vaccine to prevent this debilitating disease [2]. A systematic analysis of different vaccine candidates can result in the development of promising vaccine for human visceral leishmaniasis, more likely in the near future. Several reports have supported the agreement that sustained immunity may need constant persistence of Leishmania parasites within the host’s system [3,4]. The observation that many asymptomatic individuals mount a protective immune response after natural infection with Leishmania parasites also justifies the rationale of developing safe, whole parasite based vaccines. Diversity of host immune responses and the ability of parasites to modulate or vary their surface antigens indicate that broad antigenic repertoire in a vaccine might be essential for general human use. Considering the fact that about half of the vaccines recommended by the U.S. Food and Drug administration are developed from live or attenuated whole-cell strategy [5], it is believed that it would be beneficial to develop the technological methods for enhancing this simple approach. It is apparent that the development of clinically beneficial vaccines will involve several approaches to deliver the required safety and efficacy. Finally, questions concerning the safety and efficacy of live vaccines should be resolved and investigated before these vaccines could be examined for human use [6]. Previous reports have shown that immunization with live low dose of L. major parasites causes the differentiation of Th1-type T cell subset thereby inducing protective immunity against L. major re-infection. On the contrary, large numbers of parasites result in the expansion of Th2 cells and progressive disease [7,8]. Based on these reports, low doses of antigen inoculum have been preferred to increase the efficacy of immunization against intracellular parasites that are controlled mainly by cell mediated immunity [7]. However, it is not clear if same holds true in case of visceralizing Leishmania (L. donovani) species. Therefore, we conducted our present study to check if immunization with live low dose of L. donovani promastigotes generates protective immunity against experimental L. donovani infection in a murine model. The data generated from the present study suggests further research to develop this method as a viable approach to produce immunogenic and protective organisms for the development of effective vaccine candidates.Materials and Methods
Parasite maintenanceThe promastigotes of Leishmania donovani (MHOM/IN/80/Dd8 strain) were grown at 22±1 °C in modified NNN medium. The strain was maintained by doing serial subcultures with MEM as overlay after a regular interval of 2-3 days.
Chemicals and reagents
IgG1, IgG2a (Serotec Biorad USA), MEM (Hi-Media, Chandigarh, India), Cytokine ELISA kits (Diaclone, France), 96 well ELISA plates (Tarsons, India).
Experimental animals
Female BALB/c mice (4-5 week old) were acquired from the Central Animal House of Panjab University, Chandigarh, India. Ethical clearance for carrying out experiments was taken from Institutional Animal Ethics Committee (IAEC) of Panjab University and their guidelines of animal care and handling were followed.
Antigen preparation
Log phase promastigotes were pooled from NNN medium and centrifuged at 2500 rpm for 15 minutes. The pellet was washed 2-3 times in phosphate buffered saline (PBS). Promastigotes were enumerated in Neubauer’s chamber and a final concentration of 104 parasites per ml was prepared in sterile PBS (pH-7.2). A total volume of l00 μl containing 103 parasites was used for immunizing each animal.
Immunization protocol
Animals were divided into three groups. Immunized animals received low dose of promastigote antigen (103) in 0.1 ml of PBS via subcutaneous route. Immunized mice were challenged intravenously after three weeks with 1×107 late log phase promastigotes. Normal control animals received only PBS. Animals who received challenge infection with 107 promastigotes of Leishmania donovani served as infected controls. Different immunological parameters were investigated by euthanizing six animals in each group after immunization and three months post-infection i.e. 90 post infection/ challenge day.
Assessment of parasite load
Parasite load was estimated by studying Giemsa stained impression smears of liver. It was calculated in the form of Leishman Donovan Units (LDU) by the method of Bradley and Kirkley [9] as:
Number of amastigotes ⁄ Number of liver cell nuclei x weight of organ (in mg).
Delayed type hypersensitivity (DTH) responses
To assess the DTH responses, 48 hours before the day of sacrifice, 3 mice in each group were injected with leishmanin (40 μl) in the right and PBS (control) in the left foot pad. The thickness of both foot pads was measured after 48 hours using a pair of vernier calliper. DTH response was estimated as mean±S.D. of percentage increase in the thickness of right foot pad as compared to the left footpad of mice [10].
Antibody assay for IgG isotypes
For the estimation of IgG1 and IgG2a antibody levels serum samples were collected from animals. Conventional ELISA was carried out by the method of Ravindran et al. [11]. Briefly, 96-well ELISA plates were coated with crude antigen (5 μg/well) of L. donovani. Sera samples from all the groups were then added. All samples were run in triplicate. After that three washings were done with wash buffer and isotype specific HRP-conjugated secondary antibodies (rabbit anti-mouse IgG1 or IgG2a) were added. It was further followed by the addition of H2O2 substrate and DAB chromogen. The absorbance was calculated at 450 nm using ELISA plate reader (Lisa Plus, India).
Cytokine assays
Cytokine levels were assessed in serum samples usingcommercially available cytokine ELISA kits. The quantitative evaluation was carried out with the help of standard curve derived from the cytokine standards provided with the kit.
Statistical procedures
Data analysis was done using student’s t-test with the help of Graph Pad software. p values of less than 0.05 were considered statistically significant. Results were demonstrated as mean±S.D. of one of two independent experiments.
Results
Assessment of parasite loadTo compare the efficacy of immunization with low dose promastigotes (103) against VL, clearance of hepatic parasite burden was monitored after 90 days of challenge infection. Parasite load was quantified in the form of Leishman Donovan Units (LDU). As shown in Figure 1 mice in the infected control group exhibited maximum parasite burden and the value was 3050.16±73.92. However, in immunized group parasite load declined significantly (p<0.001) with percentage reduction of 72% as compared to infected control group and the value of LDU was 863.82±83.4.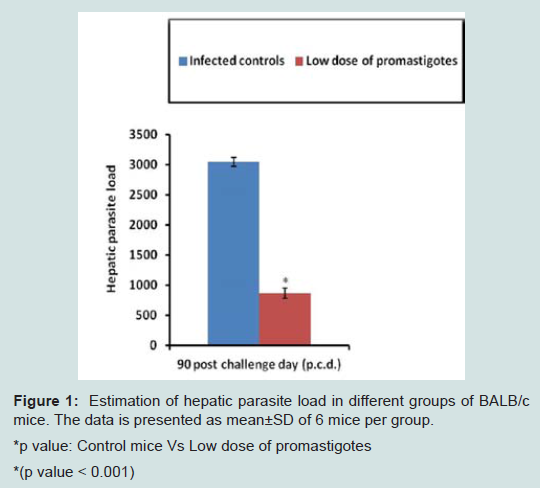
Figure 1: The amount of IgG production by vaccination. Mice were fed an AIN-93M diet and given subcutaneous vaccination at doses between 2.5 and 15.0 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HAspecific IgG antibody was measured by ELISA, as described in Materials and Methods. Values are means of 5 mice in each group. Open circles are individual data, closed circles are mean values.
DTH response was measured in immunized and control mice after 15 days of last immunization and recalled again after 90 days of challenge infection. Mice immunized with subcutaneous injection of low dose promastigote antigen displayed significant (p<0.05, p<0.001) DTH responses after immunization as well as after challenge infection in comparison to the control groups which indicates elicitation of cell mediated immunity (Figure 2).
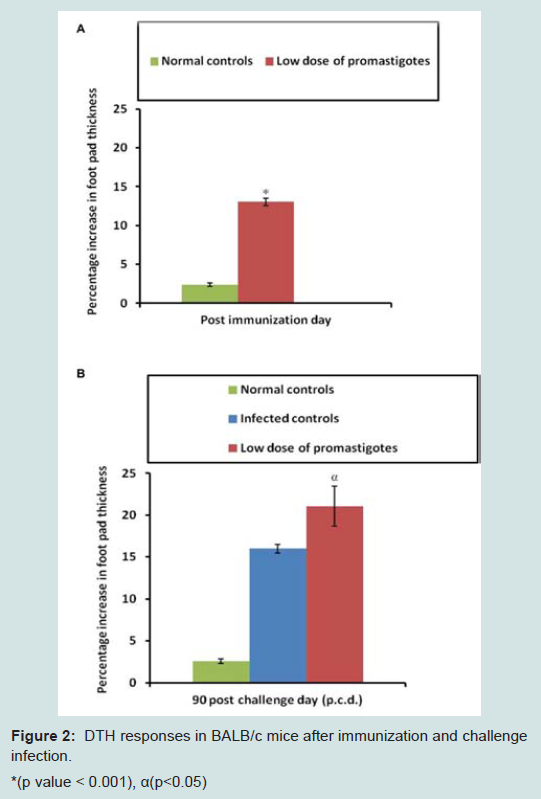
Figure 2: Effect of Lactobacillus pentosus strain S-PT84 on HA-specific IgG production in plasma. Mice were fed an AIN-93M diet with or without 0.186% S-PT84 for 6 weeks followed by subcutaneous vaccination of 2.5 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HA-specific IgG Antibody (Ab) (a) was measured by ELISA and expressed as the titer (b), as describedin Materials and Methods. Values are means of 15 mice in each group. Significant differences are seen between the control group and the S-PT84 group. Open circles are individual data, closed circles are mean values.
To evaluate the antibody levels induced by immunization, the serum levels of leishmanial antigen-specific IgG isotypes i.e. IgG1 and IgG2a were evaluated by ELISA in immunized and control groups. After immunization enhanced levels of IgG1 and IgG2a (p<0.001) were observed in the immunized group of animals in comparison to the control animals (only PBS). After challenge infection further elevation in antibody levels was noticed in the serum samples of immunized animals. However, the level of IgG2a antibody was higher as compared to IgG1 suggesting a skew towards Th1 type of immune response. On the contrary in infected control animals levels of IgG1 antibody were higher as compared to IgG2a indicating generation of Th2 type of immune response (Figure 3).
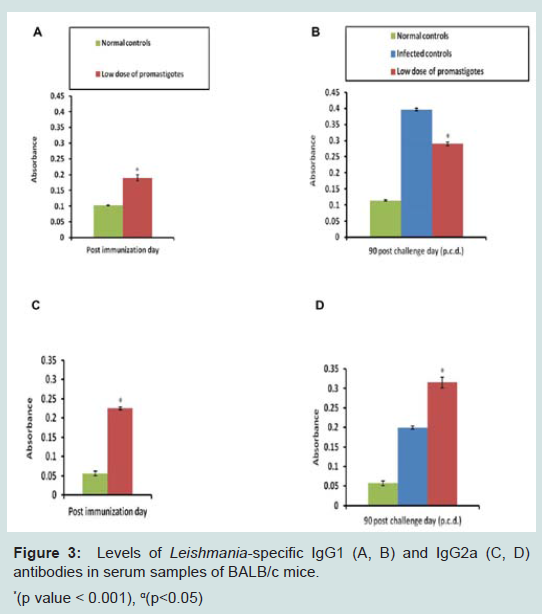
Figure 3: The amount of IgG production by vaccination. Mice were fed an AIN-93M diet and given subcutaneous vaccination at doses between 2.5 and 15.0 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HAspecific IgG antibody was measured by ELISA, as described in Materials and Methods. Values are means of 5 mice in each group. Open circles are individual data, closed circles are mean values.
A Th1 type cytokine milieu results in the effective clearance of parasites, while a Th2 cytokine type makes the host more susceptible towards infection. So, it was essential to check the levels of Th1 (IFN-γ, IL-12) and Th2 cytokines (IL-4, IL-10) in immunized and control groups. Immunized animals displayed significantly (p<0.001) higher cytokine levels after immunization over the normal controls. However, it is important to mention that level of Th1 cytokines (Figures 4 and 5) was higher over Th2 cytokines. The cytokine levels were further monitored after challenge infection. Animals of the infected control group exhibited peak levels of Th2 cytokines. Despite exhibiting substantial levels of IL-4 and IL-10 after infection, animals of the immunized group still presented high ratio of IFN-γ: IL-4 (4.95) and IFN-γ: IL-10 (1.22), possibly indicating a shift towards Th1 type of immune response (Figure 6).
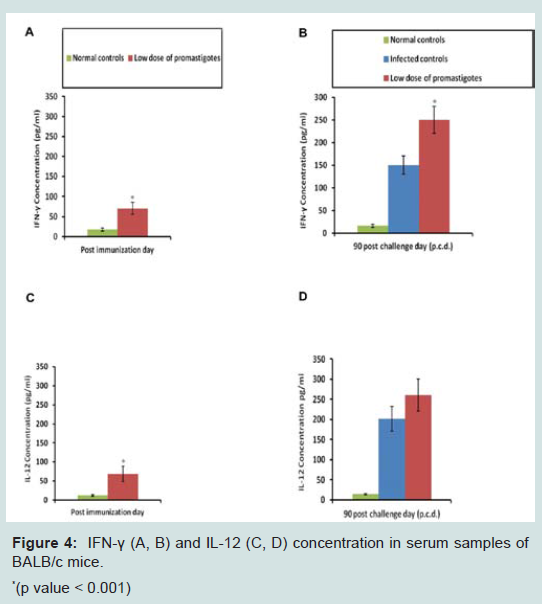
Figure 4: IFN-γ (A, B) and IL-12 (C, D) concentration in serum samples ofBALB/c mice.*(p value < 0.001)
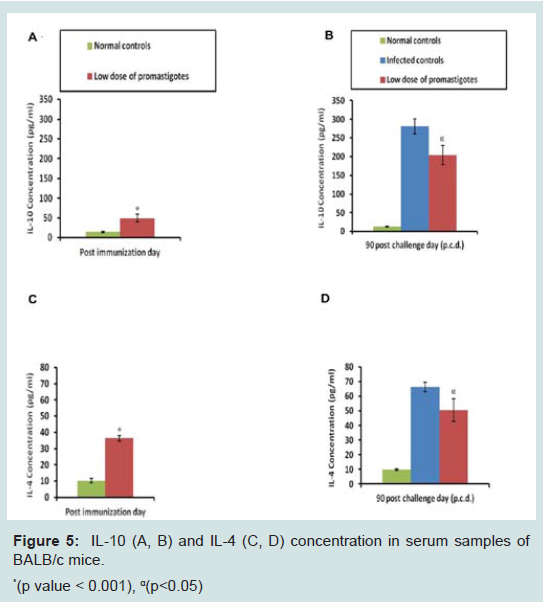
Figure 5: The amount of IgG production by vaccination. Mice were fed an AIN-93M diet and given subcutaneous vaccination at doses between 2.5 and 15.0 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HAspecific IgG antibody was measured by ELISA, as described in Materials and Methods. Values are means of 5 mice in each group. Open circles are individual data, closed circles are mean values.
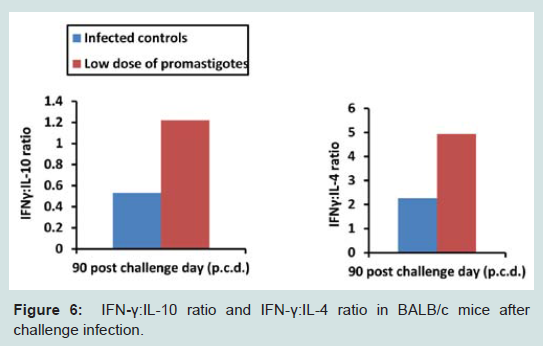
Figure 6: The amount of IgG production by vaccination. Mice were fed an AIN-93M diet and given subcutaneous vaccination at doses between 2.5 and 15.0 ng HA and boosted on day 28 from the initial vaccination. Two weeks after the last vaccination, plasma was collected from each mouse, and the amount of HAspecific IgG antibody was measured by ELISA, as described in Materials and Methods. Values are means of 5 mice in each group. Open circles are individual data, closed circles are mean values.
Discussion
Our current study is focused on L. donovani which is the causal organism of fatal disseminated visceral leishmaniasis in humans. Current treatment of the disease imposes many side effects. In addition, the development of drug resistance presents another problem in the control of this disease. In current scenario vaccination remains the driving force for the control of disease as recovery from infection confers immunity against re-infection. Thus, considerable focus has been shifted towards finding appropriate prophylactic candidates that can lead to the induction of protective immune responses against VL. Despite assessment of multiple potential candidates, no licensed vaccine is available against this disease [12]. Previous studies have shown that subclinical infection can lead to the development of cell mediated immunity as the same is noticed by a positive DTH skin test [13]. These cell mediated immune responses serve a crucial role in determining protection against re-infection. Therefore, the aim of developing anti-leishmanial vaccines is to reproduce this naturally gained protective immunity via immunizing with parasite antigens. Many studies have shown that protective cell mediated immune responses are generated in BALB/c mice immunized with small numbers of live L. major promastigotes [7,14]. This protection results from the activation of DTH responses producing Th1-type CD4+ T cells in response to low dose of antigen [8,15]. It has also been reported that dose of antigen has a profound effect on the development of T cell subsets but such reports are controversial. Some studies advocate that low-dose inoculation of parasite augments the Th1 immune response and high-dose inoculation of parasite results in the establishment of Th2 immune response, while others propose vice-versa [13]. The purpose of our study was to check whether immunization with low dose of live promastigotes can result in protection against homologous challenge infection in murine model. The protective efficacy of antigen formulation was assessed by the percentage reduction in parasite count. Similarly immunogenicity was assessed through parameters like evaluation of DTH responses, production of anti-leishmanial antibodies (IgG1 and IgG2a) and cytokines such as IFN-γ, IL-12, IL-4 and IL-10.To assess the protective efficacy of antigen formulation parasite load was assessed in infected and immunized groups on 90 post infection/challenge day. Maximum protective efficacy was observed in animals immunized with low dose of antigen which showed significant reduction in parasite load as compared to infected controls. This suggests effectiveness of this formulation in decreasing the parasite burden. This protection can be attributed to the fact that slow release of antigen from subcutaneous route allows better presentation of antigen through antigen presenting cells. Antigen is not rapidly lost from the site of injection thereby giving ample time o host immune system to generate sufficient resources to resist the subsequent infection [16]. Our results correlate very well with these ssumptions. Results are also in consistence with the study of Streit et al. where immunization and subclinical infection with live but not attenuated parasites resulted in protection against L. chagasi infection [14]. This implies that virulence of the immunizing organisms is essential to impart protective immunity.
In leishmaniasis cellular rather than humoral immune responses are involved in the recovery from disease [17,18]. Cure of VL basically relies on the acquisition of strong DTH response which results in the development of classical cell mediated immunity [19]. Corresponding to that in our study heightened foot-pad swellings were observed 48 h after inoculation of leishmanin antigen in immunized group which suggested the generation of protective immune responses. Results are similar with a study where subcutaneous immunization of mice with low-dose of L. major parasites resulted in protection against challenge infection of L. major in the footpad [14]. Also it is in accordance with the experimental studies which have shown that containment from infection is associated with strong cell-mediated DTH responses [20,21]. Since the infection is associated with impaired cell-mediated immune responses, lowest DTH responses were noticed in infected control animals. Results are similar to a study, where untreated VL patients were unable to control the infection with Leishmania donovani and it was attributed to a defective cell-mediated immune response to leishmanial antigens as these patients were unresponsive to DTH skin tests [22]. Therefore, the findings of the present study stipulate that given antigen formulation is sufficiently immunogenic and can induce significant protective cellular immune responses.
Leishmania donovani is an obligatory intracellular parasite that proliferates in macrophages. Immune recovery relies on the intracellular killing of parasite after macrophage activation via cytokines such as IL-12 and IFN-γ secreted by Th1 cells [23]. In ourstudy we estimated the cytokine profile in all the groups and observed that in case of group immunized with low dose of viable parasites, levels of Th1 cytokines (IFN-γ, IL-12) were higher over Th2 cytokines which indicates the proficiency of this dose for future vaccine studies. Generation of long-term protective immunity requires induction of memory T cells after immunization which upon encounter with the parasite can stimulate the secretion of protective Th1-like cytokines [24]. It was interesting to notice that substantial levels of IFN-γ and IL-12 were also detected after immunization which reflects priming of such memory cells, and at the time of challenge infection this memory is revoked with adequate efficacy to impart host protection. In the earlier studies, it has been well established that Th2 cytokines restricts the activation of Th1 cytokines which deactivate macrophages thereby promoting intracellular replication of parasites and exacerbation of disease [25]. Our results are in correspondence with these studies as animals of the infected control group exhibited maximum levels of these Th2 (IL-4 and IL-10) cytokines which correlated well with the higher parasite count observed in this group.
Human infection with Leishmania parasites is characterized by the presence of anti-leishmanial antibodies. Although cellmediated immunity is essentially required for the control of VL, but Leishmania-specific humoral immune responses are also detected during the infection [26,27]. In murine model, high levels of IgG1 antibody point towards Th2 type of immune response while IgG2a antibody is suggestive of Th1 type of immune response [28]. Corresponding to that in our study high levels of IgG2a antibody over IgG1 were observed in immunized animals as compared to infected controls. Our results are supported by other studies which have advocated that higher levels of IgG2a and/or lower levels of IgG1 toward promastigote antigens indicates a Th1 immune response and a disease controlling phenotype [29-31]. The potential role of antibody-mediated protection against Leishmania parasites underlines the significance of identifying those antigens that can induce the production of protective antibodies [32].
Vaccination seems to be an effective way of controlling infectious diseases. Using our subcutaneous immunization model, we conclude that protective immune responses against the visceralizing species of Leishmania can be developed by using low dose immunization regimen in mice. Moreover, for further studies newer vaccine adjuvants can be tried which can boost the effector and memory immune responses triggered by whole-cell vaccines.
References
- World Health Organization (2015) Leishmaniasis. Fact sheet No375.
- Jain K, Jain NK (2015) Vaccines for visceral leishmaniasis: a review. J Immunol Methods 422: 1-12.
- Badaro R, Jones TC, Lorenco R, Cerf BJ, Sampaio D, et al. (1986) A prospective study of visceral leishmaniasis in an endemic area of Brazil. J Infect Dis 154: 639-649.
- Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL (2002) CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 420: 502-507.
- Hoffman SL, Billingsley PF, James E, Richman A, Loyevsky M, et al. (2010) Development of a metabolically active, non-replicating sporozoite vaccine to prevent Plasmodium falciparum malaria. Hum Vaccin 6: 97-106.
- Bruhn KW, Birnbaum R, Haskell J, Vanchinathan V, Greger S, et al. (2012) Killed but metabolically active Leishmania infantum as a novel whole-cell vaccine for visceral leishmaniasis. Clin Vaccine Immunol 19: 490-498.
- Bretscher PA, Wei G, Menon JN, Bielefeldt-Ohmann H (1992) Establishment of stable, cell mediated immunity that makes “susceptible” mice resistant to Leishmania major. Science 257: 539-542.
- Doherty TM, Coffman RL (1996) Leishmania major: effect of infectious dose on T cell subset development in BALB/c mice. Exp Parasitol 84: 124-135.
- Bradley DJ, Kirkley J (1977) Regulation of Leishmania populations within the host. I. the variable course of Leishmania donovani infections in mice. Clin Exp Immunol 30: 119-129.
- Kaur S, Kaur T, Garg N, Mukherjee S, Raina P, et al. (2008) Effect of dose and route of inoculation on the generation of CD4+ Th1⁄ Th2 type of immune response in murine visceral leishmaniasis. Parasitol Res 103: 1413-1419.
- Ravindran R, Anam K, Bairagi BC, Saha B, Pramanik N, et al. (2004) Characterization of immunoglobulin G and its subclass response to Indian kala-azar infection before and after chemotherapy. Infect Immun 72: 863-870.
- Murray HW, Berman JD, Davies CR, Saravia NG (2005) Advances in leishmaniasis. Lancet 366: 1561-1577.
- Uzonna JE, Wei G, Yurkowski D, Bretscher P (2001) Immune elimination of Leishmania major in mice: implications for immune memory, vaccination, and reactivation disease. J Immunol 167: 6967-6974.
- Streit JA, Recker TJ, Filho FG, Beverley SM, Wilson ME (2001) Protective immunity against the protozoan Leishmania chagasi is induced by subclinical cutaneous infection with virulent but not avirulent organisms. J Immunol 166: 1921-1929.
- Phillips MA, Coffino P, Wang CC (1987) Cloning and sequencing of the ornithine decarboxylase gene from Trypanosoma brucei: implications for enzyme turnover and selective difluoromethylornithine inhibition. J Biol Chem 262: 8721-8727.
- Khan AA, Mudassir J, Mohtar N, Darwis Y (2013) Advanced drug delivery to the lymphatic system: lipid-based nanoformulations. Int J Nanomedicine 8: 2733-2744.
- Carvalho EM, Johnson WD, Barreto E, Marsden PD, Costa JL, et al. (1985) Cell mediated immunity in American cutaneous and mucosal leishmaniasis. J Immunol 135: 4144-4148.
- Scott P, Natovitz P, Coffman RL, Pearce E, Sher A (1988) Immunoregulation of cutaneous leishmaniasis. T cell lines that transfer protective immunity or exacerbation belong to different T helper subsets and respond to distinct parasite antigens. J Exp Med 168: 1675-1684.
- Alexander J, Carter KC, Al-Fasi N, Satoskar A, Brombacher F (2000) Endogenous IL-4 is necessary for effective drug therapy against visceral leishmanaisis. Eur J Immunol 30: 2935-2943.
- Hailu A, Menon JN, Berhe N, Gedamu L, Hassard TH, et al. (2001) Distinct immunity in patients with visceral leishmaniasis from that in subclinically infected and drug-cured people: implications for the mechanism underlying drug cure. J Infect Dis 184: 112-115.
- Banerjee A, De M, Ali N (2011) Combination therapy with paromomycin-associated stearylamine-bearing liposomes cures experimental visceral leishmaniasis through Th1-based immunomodulation. Antimicrob Agents Chemother 55: 1661-1670.
- de Andrade TM, Teixeira R, de Andrade JA, Pereira C, de Carvalho Filho EM (1982) Hypersensitivity of delayed type in visceral leishmaniasis. Rev Inst Med Trop Sao Paulo 24: 298-302.
- Sjolander A, Baldwin TM, Curtis JM, Handman E (1998) Induction of a Th1 immune response and simultaneous lack of activation of a Th2 response are required for generation of immunity to leishmaniasis. J Immunol 160: 3949-3957.
- Zaph C, Uzonna J, Beverley SM, Scott P (2004) Central memory T cells mediate long-term immunity to Leishmania major in the absence of persistent parasites. Nat Med 10: 1104-1110.
- Awasthi A, Mathur RK, Saha B (2004) Immune response to Leishmania infection. Indian J Med Res 119: 238-258.
- Miles SA, Conrad SM, Alves RG, Jeronimo SM, Mosser DM (2005) A role for IgG immune complexes during infection with the intracellular pathogen Leishmania. J Exp Med 201: 747-754.
- Mukbel R, Petersen CA, Jones DE (2006) Soluble factors from Leishmania major specific CD4+ T cells and B cells limit L. amazonensis amastigote survival within infected macrophages. Microbes Infect 8: 2547-2555.
- Ebrahimpoor S, Pakzad SR, Ajdary S (2013) IgG1 and IgG2a profile of serum antibodies to Leishmania major amastigote in BALB/c and C57BL/6 mice. Iran J Allergy Asthma Immunol 12: 361-367.
- Tewary P, Sukumaran B, Saxena S, Madhubala R (2004) Immunostimulatory oligodeoxynucleotides are potent enhancers of protective immunity in mice immunized with recombinant ORFF leishmanial antigen. Vaccine 22: 3053-3060.
- Baptista-Fernandes T, Marques C, Roos Rodrigues O, Santos-Gomes GM (2007) Intra-specific variability of virulence in Leishmania infantum zymodeme MON-1 strains. Comp Immunol Microbiol Infect Dis 30: 41-53.
- Rostamian M, Sohrabi S, Kavosifard H, Niknam HM (2015) Lower levels of IgG1 in comparison with IgG2a are associated with protective immunity against Leishmania tropica infection in BALB/c mice. J Microbiol Immunol Infect [Epub ahead of print].
- Ozbilge H, Aksoy N, Gurel MS, Yazar S (2006) IgG and IgG subclass antibodies in patients with active cutaneous leishmaniasis. J Med Microbiol 55: 1329-1331.