
Journal of Parkinsons disease and Alzheimers disease
Download PDF
Research Article
*Address for Correspondence: Clivel G. Charlton, Department of Neuroscience and Pharmacology, College of Medicine, Meharry Medical College, 1005 DB Todd Blvd. Nashville, TN 37208-3599, USA, Tel: 615 327 6510; Fax: 615 327 6632; E-mail: ccharlton@mmc.edu
Citation: Muthian G, Smith M, Dent L, King J, Griffin B, et al. Curcumin Prevents and Ameliorates Biochemical and Behavioral Toxicities of MPTP in C57BL/6J Mice: Its Potential use in Preventing and Treating Parkinsonism.. J Parkinsons Dis Alzheimer Dis. 2015;2(2): 10.
Copyright © 2015 Charlton et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Parkinson’s disease & Alzheimer’s disease | ISSN: 2376-922X | Volume: 2, Issue: 2
Submission: 05 August 2015 | Accepted: 05 October 2015 | Published: 08 October 2015
 Prenatal exposure to CU blocked the MPTP-induced reduction of mouse motor activity
Prenatal exposure to CU blocked the MPTP-induced reduction of mouse motor activity

Prenatal or embryonic (e) administration of curcumin (CUe) markedly reduced the movement-related toxicity caused by the prenatal as well as postnatal exposure to MPTP. CUe given to mice that were exposed to prenatal MPTP (CUe-MPTPe-PBS group), restored or elevated in cases of TD, all the movement parameters measured and were reduced by the exposure to prenatal MPTP, evidence in Figure 2, A, B, C and D, column 2 vs 5). Interestingly, the prenatal MPTP (MPTPe-PBS, Figure 2 column 2, Table 1 group 2) reduced TD by 42.4%, compared with the control, PBSe-PBS column 1. CUe reversed and enhanced the distance travelled (column 1 and 2 vs 5). Prenatal administered CU also reverse and enhance the toxic effect on TD caused by the combined prenatal and postnatal exposure to MPTP; evident by comparing the MPTPe-MPTP 10 group (column 4) with the CUe-MPTPe-MPTP 10 group (column 6). For MT, NM, and SC prenatal CU also reversed the toxicities caused by prenatal MPTP and the combined prenatal and postnatal MPTP (column 2 vs 5 and 4 vs 6).
 Curcumin has ameliorating effects on prenatal MPTPinduced reduction of striatal DA and its metabolites, DOPAC and 3-MT, in mice
Curcumin has ameliorating effects on prenatal MPTPinduced reduction of striatal DA and its metabolites, DOPAC and 3-MT, in mice
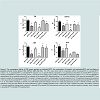
 It is of much interest that CU prevented the loss of striatal DA caused by the prenatal exposure to MPTP ( Figure 3, column 5 and 6 vs 2 and 3). When administered to prenatal-MPTP exposed offspring, CU restored DA level to 97.3% of the control ( Figure 3, PBSe-PBS, column 1 vs CUe-MPTP-PBS, column 5, as compared to the reduction to 47.2% seen in offspring exposed to prenatal-MPTP ( Table 3, group 2 and Figure 3, column 2).The HPLC analysis shows that CU also ameliorated the loss of DA that was caused by the exposure of offspring to prenatal-MPTP and postnatal-MPTP( Figure 3, CUe-MPTPe-MPTP10 group). The MPTPe-MPTP10 group, Figure 3, column 4, showed a 32.8% reduction of DA ( Table 3), whereas the pre-exposure to CU restored DA to 68.7% of the control (group 6, CUe-MPTPe-MPTP10). Table 3 also shows that the degree of changes in DA caused by the prenatal and/or postnatal exposure to MPTP and the restoration of DA levels caused by curcumin (CU) were reflected in the immediate oxidative metabolite of DA, DOPAC, and the methyl metabolite of DA, 3-MT ( Table 3). The prenatal and postnatal exposure to MPTP marked effects on the oxidative metabolite (DOPAC) and the methylated metabolite (3-MT) of DA, reducing DOPAC to 11.2% and 3-MT to 20.4% of the control. Accordingly, curcumin (CU) restores the oxidative turnoverrate (DOPAC/DA) of DA from 0.57 (10.9/19.2) in offspring exposed to prenatal and postnatal MPTP to 1.24 (59.4/47.8) in offspring that were exposed to prenatal and postnatal MPTP and received prenatal CU ( Table 2). The methylation turnover-rate of DA was likewise restored from a value of 0.16 (3.0/19.2 for the MPTPe-MPTP10 group to 0.27 (13.1/47.8) for the CUe-MPTPe-MPTP10 offspring.
It is of much interest that CU prevented the loss of striatal DA caused by the prenatal exposure to MPTP ( Figure 3, column 5 and 6 vs 2 and 3). When administered to prenatal-MPTP exposed offspring, CU restored DA level to 97.3% of the control ( Figure 3, PBSe-PBS, column 1 vs CUe-MPTP-PBS, column 5, as compared to the reduction to 47.2% seen in offspring exposed to prenatal-MPTP ( Table 3, group 2 and Figure 3, column 2).The HPLC analysis shows that CU also ameliorated the loss of DA that was caused by the exposure of offspring to prenatal-MPTP and postnatal-MPTP( Figure 3, CUe-MPTPe-MPTP10 group). The MPTPe-MPTP10 group, Figure 3, column 4, showed a 32.8% reduction of DA ( Table 3), whereas the pre-exposure to CU restored DA to 68.7% of the control (group 6, CUe-MPTPe-MPTP10). Table 3 also shows that the degree of changes in DA caused by the prenatal and/or postnatal exposure to MPTP and the restoration of DA levels caused by curcumin (CU) were reflected in the immediate oxidative metabolite of DA, DOPAC, and the methyl metabolite of DA, 3-MT ( Table 3). The prenatal and postnatal exposure to MPTP marked effects on the oxidative metabolite (DOPAC) and the methylated metabolite (3-MT) of DA, reducing DOPAC to 11.2% and 3-MT to 20.4% of the control. Accordingly, curcumin (CU) restores the oxidative turnoverrate (DOPAC/DA) of DA from 0.57 (10.9/19.2) in offspring exposed to prenatal and postnatal MPTP to 1.24 (59.4/47.8) in offspring that were exposed to prenatal and postnatal MPTP and received prenatal CU ( Table 2). The methylation turnover-rate of DA was likewise restored from a value of 0.16 (3.0/19.2 for the MPTPe-MPTP10 group to 0.27 (13.1/47.8) for the CUe-MPTPe-MPTP10 offspring.
 In addition to striatal TH, midbrain TH was also studied. As shown in Figure 5, the main difference between the changes in striatal and midbrain TH is that the midbrain TH level was significantly decreased by the postnatal exposure to MPTP ( Figure 5, lane 2 vs 1 in A; 2nd column vs the 1st column in B). CU (15 μg/animal) administered to offspring exposed to prenatal MPTP and to offspring exposed to both prenatal and postnatal MPTP enhancing the expression of TH ( Figure 5, lanes 5 and 6).
In addition to striatal TH, midbrain TH was also studied. As shown in Figure 5, the main difference between the changes in striatal and midbrain TH is that the midbrain TH level was significantly decreased by the postnatal exposure to MPTP ( Figure 5, lane 2 vs 1 in A; 2nd column vs the 1st column in B). CU (15 μg/animal) administered to offspring exposed to prenatal MPTP and to offspring exposed to both prenatal and postnatal MPTP enhancing the expression of TH ( Figure 5, lanes 5 and 6).
Curcumin Prevents and Ameliorates Biochemical and Behavioral Toxicities of MPTP in C57BL/6J Mice: Its Potential use in Preventing and Treating Parkinsonism
Gladson Muthian, Marquitta Smith, Lemuel Dent, Jennifer King, Brenya Griffin, Veronica Mackey and Clivel Charlton*
- Department of Neuroscience and Pharmacology, Meharry Medical College, Nashville, USA
*Address for Correspondence: Clivel G. Charlton, Department of Neuroscience and Pharmacology, College of Medicine, Meharry Medical College, 1005 DB Todd Blvd. Nashville, TN 37208-3599, USA, Tel: 615 327 6510; Fax: 615 327 6632; E-mail: ccharlton@mmc.edu
Citation: Muthian G, Smith M, Dent L, King J, Griffin B, et al. Curcumin Prevents and Ameliorates Biochemical and Behavioral Toxicities of MPTP in C57BL/6J Mice: Its Potential use in Preventing and Treating Parkinsonism.. J Parkinsons Dis Alzheimer Dis. 2015;2(2): 10.
Copyright © 2015 Charlton et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Parkinson’s disease & Alzheimer’s disease | ISSN: 2376-922X | Volume: 2, Issue: 2
Submission: 05 August 2015 | Accepted: 05 October 2015 | Published: 08 October 2015
Abstract
Parkinson’s disease (PD) is the result of the degeneration of nigrostriatal dopamine neurons and loss of striatal tyrosine hydroxylase and dopamine. The cause is unknown. Studies have shown that fetal exposures to toxins make the nigrostriatal dopamine neurons vulnerable and that later toxic challenges and/or the wear-and-tear of aging cause the already vulnerable neurons to succumb, and producing a model of parkinsonism. Thus, a sensitization stage anda precipitating stage may exist for idiopathic PD. In this study, we used 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) to produce the proposed sensitization and precipitating stages and determine if curcumin can ameliorate the toxic changes. Note: however that MPTP serves as a protoxin, via it metabolism by monoamine axidase B (MAO-B) to 1-methyl-4-phenylpyridinium (MPP+). MPP+ has affinity for the catecholamine uptake system, is taken up preferentially into dopaminergic neurons with cell bodies in the substantia nigra zona compacta, where it inhibits complex-1 of the electron transport chain. C57BL/6J pregnant mice were treated with low dosage of MPTP during gestation days (GD) 8-12, to cause sub-threshold impairments to the emerging and vulnerable nigrostriatal dopamine neurons in the fetus, thus producing phenotypic sensitization. At 3 months, the offspring of the PBS control and the MPTP group were challenged with PBS or MPTP (10 mg/kg/day) for seven days, to cause further harm and to the produce the precipitating stage. The ameliorating effect of curcumim was tested on both stages, by pretreating with 15 μg/animal during GD 3-12. All animals were studied for motor activities, the levels of TH, and dopamine and its metabolites. The prenatal treatments with curcumin nullify the reduction in birth weight caused by prenatal MPTP, and it blocked the impairments in movement caused by prenatal MPTP, postnatal MPTP and combination of prenatal and postnatal MPTP. Curcumin also restored dopamine to 97.3% from the 47.2% loss caused by prenatal MPTP, and to 68.7% from the 32.8% loss caused by the prenatal and postnatal MPTP treatments. TH expression was also markedly restored. These novel data show the efficacy of curcumin and suggest that the curcumin may be valuable in preventing and ameliorate PD-like changes.Keywords
Curcumin; Dopamine; Locomotor activity; MPTP; Tyrosine hydroxylase; Parkinson’s diseaseAbbreviations
CU: Curcumin; Cue: Curcumin Embryonic Exposed; DA: Dopamine; DOPAC: 3,4-Dihydroxyphenylacetic acid; HVA: Homo Vanillic Acid; MPTP: 1 methyl-4-phenyl-1,2,3,6-tetrahydropyridine; MT: Movement Time; NM: Number of Movements; NS: Nigrostriatum; PD: Parkinson’s Disease; TD: Total Distance; TH: Tyrosine Hydroxylase; 3-MT: 3-MethoxytyramineIntroduction
Parkinson’s (PD) disease is an age-related neurodegenerative disease caused by the loss of dopaminergic neurons within the substantianigra (SN). This results in extrapyramidal motor dysfunction, including tremor, rigidity, and bradykinesia [1] and decreased tyrosine hydroxylase (TH) and the production and storage of dopamine (DA) in the striatum [2,3]. At present, DA replacement therapy with levo-dopa, or other drugs with dopaminergic agonists’ properties, can alleviate the symptoms of the disorder. However, although such drugs are effective in the early stages of the disease, long- term therapy has been associated with serious side effects.Oxidative stress and mitochondrial dysfunction are major events that occur during neuronal death that causes PD [4,5]. This has been noted as glutathione (GSH) depletion in dopaminergic cells and mitochondrial dysfunction [6]. Thus, the therapeutic approach of PD treatment could include not only the amelioration of the DA loss, but also the modulation of oxidative stress. Phenolic antioxidants, including flavonoids are an extensive group of naturally-occurring compounds that are widely distributed in plants, and are constituents of various fruits, nuts and leaves [7,8]. Flavonoids are potent antioxidants and free radical scavengers, with efficacy suggested to exceed the antioxidant capacity of vitamins C and E [9-12]. Flavonoids are capable of chelating metal ions and modifying the activity of cellular antioxidants and antioxidant enzymes, such as catalase and GSH [13]. The flavonoids also modulate nitric oxide production, tumor necrosis factor alpha secretion and nuclear factor kβ (NFkβ) dependent gene expression, in vitro, [14] and have anti-inflammatory properties [15-19]. These agents also inhibit the activities of lipoxygenase and cycloxygenase. Moreover, as model antioxidants, they show very little toxicity, even in long-term studies, and they possibly slow down the rate of nigral cell loss in PD [20].
Curcumin (CU) is a well-known food flavor, a naturally-occurring yellow pigment isolated from the rhizomes of the plant found in South Asia [21,22]. It is a diarylheptanoid polyphenol isolated from the rhizomes of, and is abundant in, turmeric (Curcuma longa), a plant that has been used in India for centuries as a food preservative and medicinal agent for the treatment of human disease associated with injury and inflammation [23,24], for neurological disorders, such as Alzheimer’s disease (AD) and for symptoms associated with stroke [25,26]. It has been shown that curcumin (CU) protects PC12 cells against 1-methyl-4-phenylpyridinium (MPP+) neurotoxicity, through its anti-apoptotic and antioxidative properties [27]. It has also been suggested that curcumin crosses the blood-brain barrier and enters the central nervous system [28], which supports the possibility that CU might be an effective therapeutic agent for neurodegenerative disorders.
1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) has been shown to cause the syndrome in humans that resemble PD, and have been well accepted as a neurotoxin and produces a reliable model of PD [29,30]. Moreover, low dosage of MPTP, which does not affect the pregnant dams, has been shown to cause subtle prenatal toxicity in the offspring. When the adult offspring, that were other-wise physically normal-looking, were challenged with a second low dosage of MPTP, the effects of MPTP were additive, as shown by severe changes in markers of PD, such as TH depletion, dopamine loss and hypokinesia [31]. Such a study indicated that some cases of PD may have a fetal basis, in which subtle prenatal nigrostriatal impairments occurred-as shown by the prenatal toxicity of MPTP- and PD symptoms are precipitated later by deteriorating changes in the nigrostriatum -as shown by the postnatal toxicity of the low dosage of MPTP- that did not affect the normal prenatal control mice [31,32].
When the neurotoxin MPTP is used, it is converted to 1-methyl-4-phenylpyridinium (MPP+) by brain monoamine oxidase B (MAO-B). MPP+ is then transported into dopaminergic neurons via the DA transporter (DAT) and accumulates in the mitochondria causing oxidative damage by disrupting respiratory enzymes and causing cell death [33]. Since MPTP metabolism causes oxidative damage and results in PD-like changes, it is quite plausible that CU, serving as an antioxidant, will offer protection from the PD-like toxicity caused by MPTP. Therefore, we designed a study that examines the prenatal effect of CU on MPTP-induced toxicity in a mouse model of PD. The model used consists of C57BL/6J mice, which were exposed inutero to low dosage of MPTP or to PBS during the critical period of the birth of the NS DA neurons [31]. The MPTP exposed offspring showed subtle impairments of the basal ganglia DA system but with no visual toxicity, but when they were exposed to low dosage of postnatal-MPTP they exhibited the level of changes equated to those that occur in PD-like. Thus, the prenatal MPTP exposures produce a sensitization stage for PD and the postnatal exposure to MPTP represents the precipitation stage. The effects of CU were tested in an MPTP-sensitization group of offspring as well as in offspring, which were exposed to the precipitating dosage of MPTP. Interestingly, CU showed ameliorating effects on the toxicity caused by MPTP.
Materials and Methods
ChemicalsCurcumin (1,7-bis-4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) was purchased from Calbiochem, La Jolla, CA. Polyclonal TH antibody was purchased from Chemicon, USA. MPTP was purchased from Sigma Chemical Co., St Louis, MO USA.
Experimental design
Animals: Young adult pregnant (eight weeks old) C57BL/6J mice were purchased from Jackson Laboratory, Bar Harbor, ME. The dams were shipped on day one of pregnancy. They were housed two per cage, under a 12-hr light-dark cycle in a 22 ºC temperature-controlled room. Pregnant dams and offspring had access to standard chow and water, ad libitum. On day 3 of arrival the animals were treated. All procedures were conducted in accordance with the Guide for Care and Use of Laboratory Animals (National Institutes of Health) and were approved (protocol # 050224CGC052-02) by the Institutional Animal Care and Use Committee (IACUC) at Meharry Medical College.
Previous studies showed that the prenatal effects of MPTP caused the nigrostriatal (NS) dopaminergic system to be sensitive to toxic or stressful type of manipulation, e.g. to low doses of MPTP administered later in life 31,32]. So, in this project, we examined the possible protective effects of curcumim (CU) on NS dopaminergic toxicity in mice exposed to prenatal MPTP, postnatal MPTP and to prenatal and postnatal MPTP. For the 1st study we examined the prenatal effects of a low toxic dose of MPTP and we determined whether postnatal MPTP exposure has an additive toxic effect with the prenatal exposure, on NS dopaminergic markers. To do so, two groups of mice were treated with PBS (1 ml/100 g), as control, or with MPTP (10 mg/kg) during gestation day 8-12, a period corresponds with the period of birth for the NS dopamine neurons. After delivery, the pups were cross-nurtured, and their weights determined. At 28 days, the pups were weaned and separated according to sex and their weights recorded until the 12th weeks of age. At 12 weeks the young adult male pups of the prenatal PBS and prenatal MPTP treated dams were randomly divided into two groups each and were treated with either PBS or MPTP (10 mg /kg. i.p.) for 7 days. Seven days after the last injection the motor activity of the mice was determined. A sub-group was sacrificed by decapitation, their brains dissected and the striatum and midbrain were used for Western blot analysis for TH, and the striatum used for the high-performance liquid chromatographic (HPLC) determination of dopamine and its metabolites, 3, 4-dihydroxyphenylacetic acid (DOPAC), 3-methoxytyramine (3-MT) and homovanillic acid (HVA).
This protocol produced 4 groups, based on the prenatal exposure to PBS or MPTP and postnatal exposure to PBS and MPTP. To illustrate, the letter ‘e’ is used to represents embryonic or fetal exposure and its absence the postnatal exposure. The comparative groups are: PBSe-PBS group vs. MPTPe-PBS group that allows us to compare the effects of the prenatal exposure to MPTP vs. PBS. The PBSe-MPTP vs. MPTPe-MPTP, which helps us to determine whether the exposure to MPTP later in life has an additive effect in animals exposed in utero to MPTPe. To study the protective effects of CU from prenatal MPTP, the fetuses were pre-exposed, at E3-12, to CUe (15 μg/animal) and MPTPe, 10 mg/kg, was given at E8-12 to the same animals. This group was then divided into two subgroups, and the 3 months old offspring were treated with either PBS or MPTP, as above. Thus, this protocol produced two groups: CUe-MPTPe-PBS vs. CUe-MPTPe-MPTP. The CUe-MPTPe-PBS group determines if CUe has a protective effect on the toxicity caused by the prenatal exposure to MPTPe, by comparing the PBSe-MPTPe-PBS vs. the CUe-MPTPe-PBS groups. By comparing the PBSe-MPTPe-MPTP vs. CUe-MPTPe-MPTP, the treatments help to show whether CU offers protection from the toxicities caused by the combined prenatal and postnatal MPTP.
Measurement of locomotor activities: Seven days after the last injection of PBS or MPTP the mice were studied for their locomotor activities. Each mouse was placed in the rodent activity monitor (Versamax Analyzer, Accusccan Instruments Inc., Columbus, OH, USA), and measurements were taken starting after three mins and continued for 10 min. The measurements were performed in a quiet isolated room with dim lighting. Movement time (MT), total distance (TD) and the number of movements (NM) and stereotypy counts (SC) were measured.
HPLC-electrochemical (EC) determination of dopamine and its metabolites: The isolated striatal tissues were homogenized in 750 μl of 0.1 M trichloro-acetic acid (TCA), which contains 10-2 M sodium acetate, 10-4 M EDTA and 10.5% methanol (pH 3.8) using a tissue dismembrator (Fisher Scientific). Samples were spun in a micro centrifuge at 10,000x g for 20 mins. Samples of the supernatant were then analyzed for dopamine and its metabolites at the core analytical facility at Vanderbilt University. Dopamine and its metabolites were determined by specific HPLC assay utilizing an Antec Decade 11 (oxidation: 0.5) electrochemical detector operated at 33 ºC. Supernatant samples of 20 μl were injected, using Water’s 717+ auto sampler, onto Phenomena Nucleosol (5 U, 100 A) C18 HPLC column (150x4.60 mm). Biogenic amines were eluted with a mobile phase consisting of 89.5% 0.1 M TCA, 10-2 M sodium acetate, 10-4 M EDTA and 10.5% methanol (pH 3.8). Solvent is delivered at 0.8 ml/min using Water’s 515 HPLC pump. Using this HPLC solvent the following biogenic amines elute in the following order: noradrenaline, adrenaline, di-hydroxyphenylacetic acid (DOPAC), dopamine (DA), 5-hydroxyindole acetic acid (5-HIAA), 5-hydroxytryptamine (5-HT), and 3-methoxytyramine (3-MT). HPLC control and data acquisition were managed by Water’s Empower software, Milford, MA.
Western blot analysis of TH protein expression: Immunoblotting was used to quantify the amount of TH and LAAD proteins in the midbrain. After decapitation, the brains were dissected and the sections were homogenized in lysis buffer (Ambion, Austin, TX, USA). Protein concentration was determined using Bio-Rad protein reagent (Bio-Rad, USA). The proteins were precipitated by adding 100% methanol and centrifuged at 10,000 rpm for 10 mins in a Sorvall refrigerated centrifuge. Then the supernatants were decanted, and the precipitates washed with 90% methanol for 10 mins. Proteins were dissolved in Lamelli sample buffer and transferred to nitrocellulose membrane. The membranes were blocked in 5% BSA for one hour. Membranes were incubated with primary rabbit TH (1: 2,000, Chemicon International, CA, USA) and then exposed to the secondary antibody 1: 10,000 (HRP conjugated anti-rabbit IgG, Sigma Chemical Co, Saint Louis, MO) and visualized by chemiluminescence. All the experiments were repeated three times to confirm the results.
Statistical analysis
All data are reported as mean ± SEM and analyzed by oneway analysis of variance (ANOVA) followed by Turkey’s multiple comparison test. P values less than 0.05 were considered significant. (PRISM, Graph pad software Inc), La Jolla CA.
Results
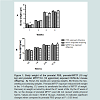
Prenatal exposure to MPTP and the effect of CU on the body weight of miceThe body weights of pups were measured and recorded once weekly before weaning and every two weeks after weaning up to 12 weeks of age. This was done to determine if in utero drug exposures cause toxic effects expressed as changes in body weights and growth of the offspring. During the 1st week of life it can be seen that the offspring exposed to 10 mg/kg MPTP showed 33% reduction in body weight as compared to those exposed to PBS ( Figure 1A, week 1). CU (3rd column for wks 1, 2, 3 and 4) partially prevented the weight loss as compared to the group exposed to prenatal PBS ( Figure 1A, 1st column for wks 1, 2, 3 and 4). The body weight of the CUe-MPTPe-offspring caught-up to the PBS group by week 4 ( Figure 1A, 2nd vs 1st columns). For week 1, 2, 3 and 4 the average body weights for the prenatal-PBS; MPTP and prenatal CU-MPTP offspring were: 1.73; 1.2; 1.52 gm; 3.30; 2.80; 3.30 gm; 6.80; 5.0; 6.40 gm and 7.32; 6.75; 7.48 gm, producing a aggregate weight for the respective groups of 19.15, 15.75 and 18.70 gm. So, prenatal MPTP exposure reduced the aggregate 4 weeks mouse weight to 82.25% of the PBS control and curcumin (CU) restored the weight to 97.65%. The body weights were not significantly different among the groups for the post-weaning 5-12 weeks period ( Figure 1B). In summary, the results show that the exposure of the fetus to prenatal MPTP during G8-G12 resulted in offspring with reduced birth weights. The MPTP exposure, however, may not affect the nurturing and growth processes, because the body weights caught up with the weight of the PBS control by the 5th week of life. CU showed a blocking/ameliorating effect on the weight reducing effects of MPTP, evident during the 1st 4 weeks.
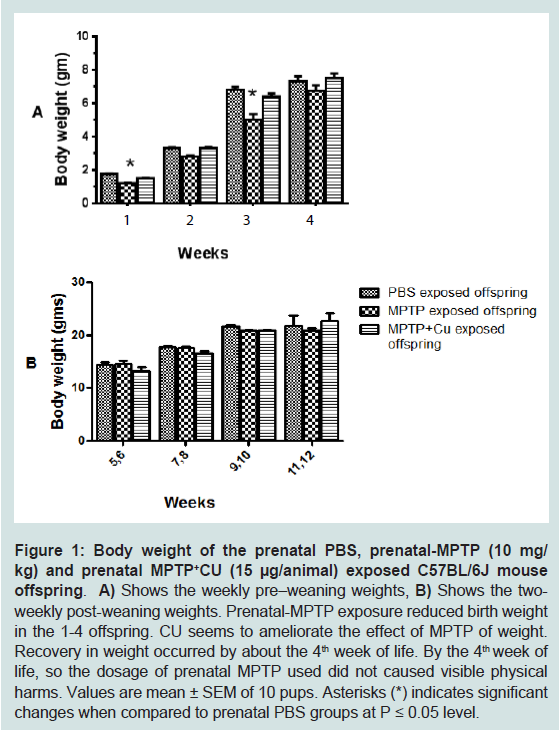
Figure 1: Body weight of the prenatal PBS, prenatal-MPTP (10 mg/kg) and prenatal MPTP+CU (15 μg/animal) exposed C57BL/6J mouse offspring.A) Shows the weekly pre–weaning weights, B) Shows the two weekly post-weaning weights. Prenatal-MPTP exposure reduced birth weight in the 1-4 offspring. CU seems to ameliorate the effect of MPTP of weight. Recovery in weight occurred by about the 4th week of life. By the 4th week of life, so the dosage of prenatal MPTP used did not caused visible physical harms. Values are mean ± SEM of 10 pups. Asterisks (*) indicates significant changes when compared to prenatal PBS groups at P ≤ 0.05 level.
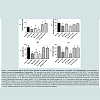
The spontaneous motor activities of mouse offspring exposed to PBS, MPTP or CU+MPTP during the fetal or embryonic stage of life were measured at 12 weeks of age. The parameters measured were the total distance travel (TD), the number of movements (NM), movement time (MT) and stereotype movement counts (SC). As shown in Figure 2, A, B, C and D, PBSe-PBS (column 1) vs MPTPe- PBS) and Table 1 group 1 vs 2, for all measures, the exposure to prenatal MPTP (MPTPe), as compared to prenatal PBS (PBSe), reduced TD, NM, MT and SC by 42.4%, 33.8%, 55.5% and 50.3% ( Figure 2, Table 1 line 1 vs 2). The same dose of 10 mg/kg MPTP given at postnatal 12 weeks of age was less effective; reducing TD, NM, MT and SC by 25.1%, 12.7%, 42.9% and 8.1% ( Figure 2, column 1 vs 3), however the combined in-utero/embryomic and post-natal MPTP (MPTPe-MPTP10, Figure 2, column 1 vs 4 for A, B, C and D) showed severe reduction in TD, NM, MT and SC. The data show that the postnatal exposure to MPTP showed additive effects with the prenatal exposure; reducing TD, NM, MT and SC by 46.6%, 36.8%, 63.1% and 64.1%. This means that postnatal MPTP enhanced the toxic effects of the prenatal MPTP by 21.5%, 24.1%, 20.2% and 55.9% for TD, NM, MT and SC.
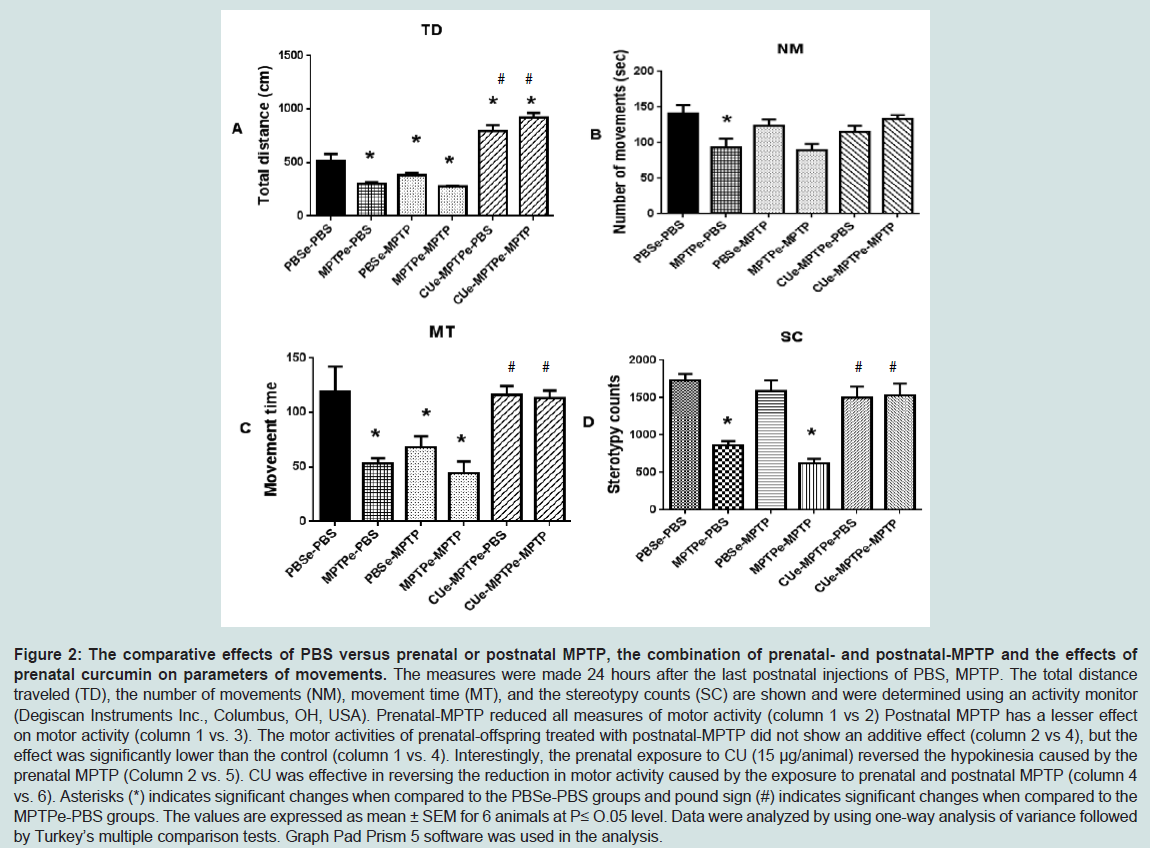
Figure 2: The comparative effects of PBS versus prenatal or postnatal MPTP, the combination of prenatal- and postnatal-MPTP and the effects of prenatal curcumin on parameters of movements. The measures were made 24 hours after the last postnatal injections of PBS, MPTP. The total distance traveled TD), the number of movements (NM), movement time (MT), and the stereotypy counts (SC) are shown and were determined using an activity monitor (Degiscan Instruments Inc., Columbus, OH, USA). Prenatal-MPTP reduced all measures of motor activity (column 1 vs 2) Postnatal MPTP has a lesser effect on motor activity (column 1 vs. 3). The motor activities of prenatal-offspring treated with postnatal-MPTP did not show an additive effect (column 2 vs 4), but the effect was significantly lower than the control (column 1 vs. 4). Interestingly, the prenatal exposure to CU (15 μg/animal) reversed the hypokinesia caused by the prenatal MPTP (Column 2 vs. 5). CU was effective in reversing the reduction in motor activity caused by the exposure to prenatal and postnatal MPTP (column 4 vs. 6). Asterisks (*) indicates significant changes when compared to the PBSe-PBS groups and pound sign (#) indicates significant changes when compared to the MPTPe-PBS groups. The values are expressed as mean ± SEM for 6 animals at P≤ O.05 level. Data were analyzed by using one-way analysis of variance followed by Turkey’s multiple comparison tests. Graph Pad Prism 5 software was used in the analysis.
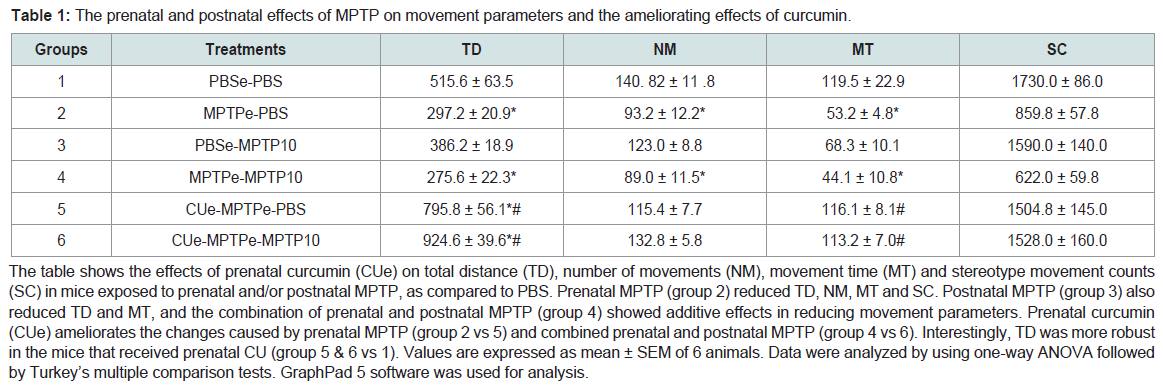
Table 1: The prenatal and postnatal effects of MPTP on movement parameters and the ameliorating effects of curcumin.
The table shows the effects of prenatal curcumin (CUe) on total distance (TD), number of movements (NM), movement time (MT) and stereotype movement counts (SC) in mice exposed to prenatal and/or postnatal MPTP, as compared to PBS. Prenatal MPTP (group 2) reduced TD, NM, MT and SC. Postnatal MPTP (group 3) also reduced TD and MT, and the combination of prenatal and postnatal MPTP (group 4) showed additive effects in reducing movement parameters. Prenatal curcumin (CUe) ameliorates the changes caused by prenatal MPTP (group 2 vs 5) and combined prenatal and postnatal MPTP (group 4 vs 6). Interestingly, TD was more robust in the mice that received prenatal CU (group 5 & 6 vs 1). Values are expressed as mean ± SEM of 6 animals. Data were analyzed by using one-way ANOVA followed by Turkey’s multiple comparison tests. GraphPad 5 software was used for analysis.
The outcome clearly demonstrates that CU protects the offspring from the prenatal toxicity effects of MPTP on movement parameters, and CU also protect offspring that were exposed to prenatal and postnatal MPTP (MPTPe-MPTP). CU ameliorated the reduction caused by prenatal MPTP from a 53.4% reduction in TD to an overall gain of 25.9% and rescue the loss in TD caused by combined prenatal and postnatal MPTP from a loss of 46.5% to a gain of 88.8%, as shown in Table 2 giving a permutation of the effects of MPTP and CU across groups. The outcomes suggest that CU has a preventative as well as an ameliorating effect on the hypokinetic type of toxicites caused by the prenatal exposure to MPTP as well as the prenatal plus postnatal exposures to MPTP ( Table 2).
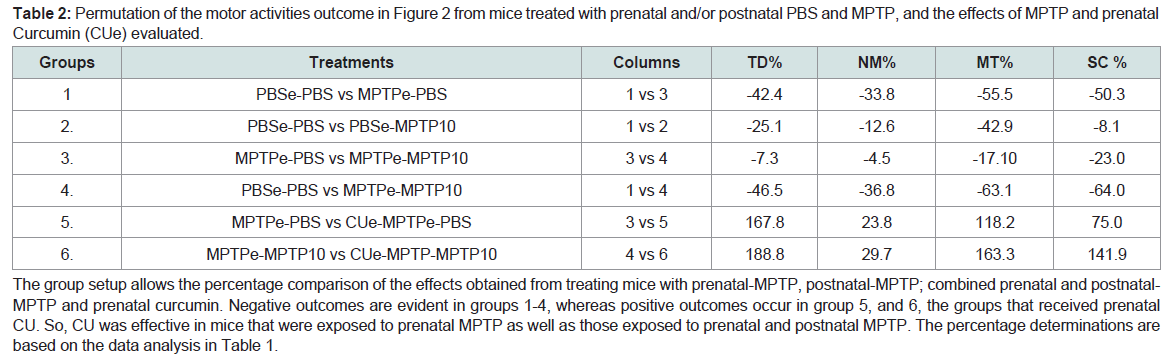
Table 2: Permutation of the motor activities outcome in Figure 2 from mice treated with prenatal and/or postnatal PBS and MPTP, and the effects of MPTP and prenatal Curcumin (CUe) evaluated.The group setup allows the percentage comparison of the effects obtained from treating mice with prenatal-MPTP, postnatal-MPTP; combined prenatal and postnatal-MPTP and prenatal curcumin. Negative outcomes are evident in groups 1-4, whereas positive outcomes occur in group 5, and 6, the groups that received prenatal CU. So, CU was effective in mice that were exposed to prenatal MPTP as well as those exposed to prenatal and postnatal MPTP. The percentage determinations are based on the data analysis in Table 1.
The concentration of dopamine (DA), 3,4-dihydroxy-phenylacetic acid (DOPAC), 3-methoxytyramine (3-MT) and homovanillic acid (HVA) were measured in the striatum of C57BL/6J mouse offspring exposed to MPTP. One group of the C57BL/6J offspring was exposed in utero at G8-12 to 10 mg/kg MPTP. Another group was exposed to the prenatal MPTP and to postnatal 10 mg/kg MPTP at 3 months of age. The control mice received PBS. Curcumin (CU), at 15 μg/animal, was administered at G3-12 to a group of prenatal MPTP (MPTPe-PBS) and a group of prenatal MPTP mice that also received postnatal MPTP treatment (MPTPe-PMTP10), to determine if CU can prevent and ameliorate the toxic effects of MPTP. As shown in Figure 3, Column 1 vs 2, and Table 3, group 2 vs1), the prenatal exposure to MPTP reduced the level of DA by 52.8%. The postnatal administration of 10 mg/kg MPTP (PBSe-MPTP10, group 3) also reduced DA to 20% of the control value ( Table 3, column 3 vs 1). Moreover, the offspring those were exposed to prenatal MPTP and also received 10 mg/kg postnatal MPTP (MPTPe-MPTP10, group 4) showed a reduction in DA of 67.2%. This is indicative of an additive effect when compared to the individual value for prenatal MPTP ( Table 3, group 2) and postnatal MPTP ( Table 3, group 3). This represents an additive toxic effect of 14.4% caused by challenging the prenatal MPTP mice with postnatal MPTP ( Figure 3A).
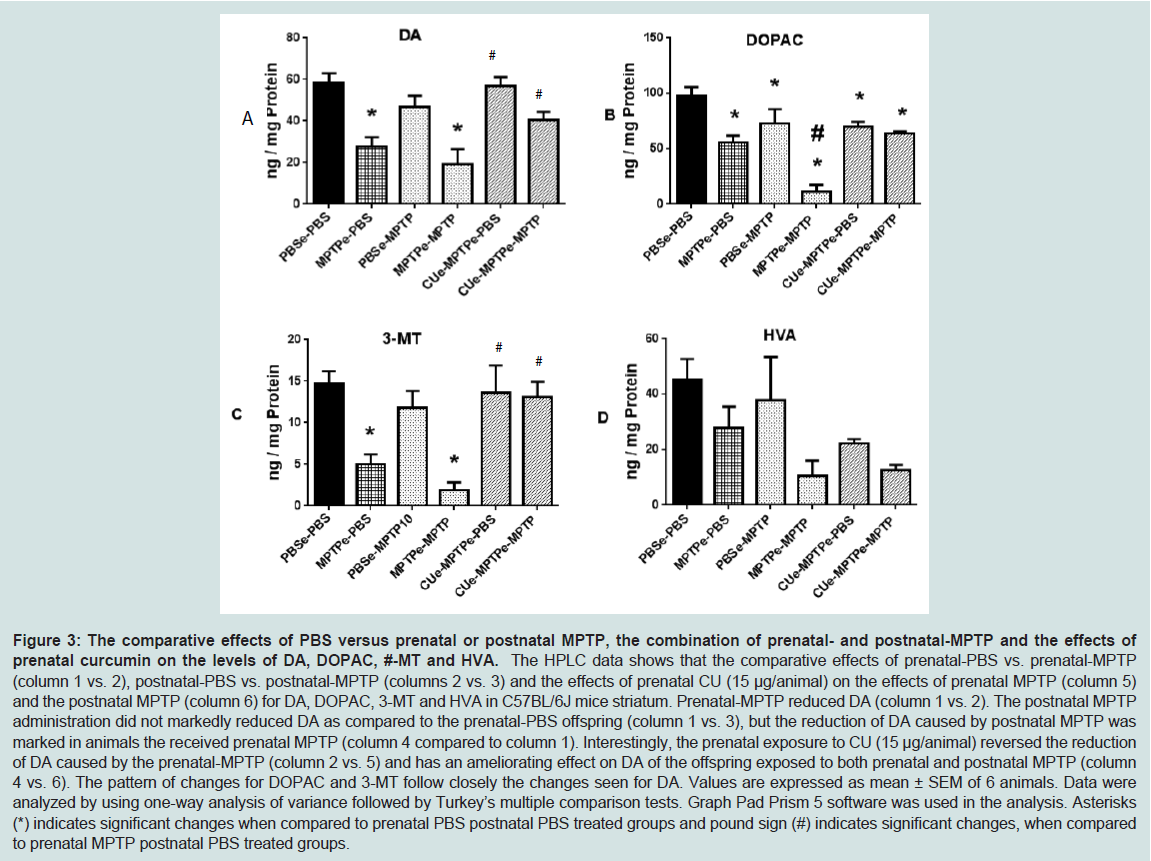
Figure 3: The comparative effects of PBS versus prenatal or postnatal MPTP, the combination of prenatal- and postnatal-MPTP and the effects of prenatal curcumin on the levels of DA, DOPAC, #-MT and HVA.The HPLC data shows that the comparative effects of prenatal-PBS vs. prenatal-MPTP (column 1 vs. 2), postnatal- BS vs. postnatal-MPTP (columns 2 vs. 3) and the effects of prenatal CU (15 μg/animal) on the effects of prenatal PTP (column 5) and the postnatal MPTP (column 6) for DA, DOPAC, 3-MT and HVA in C57BL/6J mice striatum. Prenatal-MPTP reduced DA (column 1 vs. 2). The postnatal MPTP administration did not markedly reduced DA as compared to the prenatal-PBS offspring (column 1 vs. 3), but the reduction of DA caused by postnatal MPTP was marked in animals the received prenatal MPTP (column 4 compared to column 1). Interestingly, the prenatal exposure to CU (15 μg/animal) reversed the reduction of DA caused by the prenatal-MPTP (column 2 vs. 5) and has an ameliorating effect on DA of the offspring exposed to both prenatal and postnatal MPTP (column 4 vs. 6). The pattern of changes for DOPAC and 3-MT follow closely the changes seen for DA. Values are expressed as mean ± SEM of 6 animals. Data were analyzed by using one-way analysis of variance followed by Turkey’s multiple comparison tests. Graph Pad Prism 5 software was used in the analysis. Asterisks (*) indicates significant changes when compared to prenatal PBS postnatal PBS treated groups and pound sign (#) indicates significant changes, when comparedto prenatal MPTP postnatal PBS treated groups.
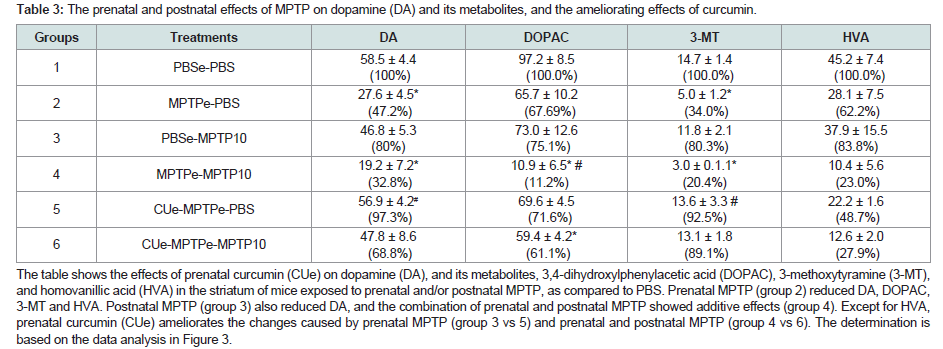
Table 3: The prenatal and postnatal effects of MPTP on dopamine (DA) and its metabolites, and the ameliorating effects of curcumin.The table shows the effects of prenatal curcumin (CUe) on dopamine (DA), and its metabolites, 3,4 dihydroxylphenylacetic acid (DOPAC), 3-methoxytyramine (3-MT), and homovanillic acid (HVA) in the striatum of mice exposed to prenatal and/or postnatal MPTP, as compared to PBS. Prenatal MPTP (group 2) reduced DA, DOPAC, 3-MT and HVA. Postnatal MPTP (group 3) also reduced DA, and the combination of prenatal and postnatal MPTP showed additive effects (group 4). Except for HVA, prenatal curcumin (CUe) ameliorates the changes caused by prenatal MPTP (group 3 vs 5) and prenatal and postnatal MPTP (group 4 vs 6). The determination is based on the data analysis in Figure 3.
Prenatal CU blocked the reduction of the expression of striatal TH caused by MPTP
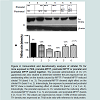
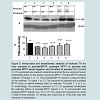
The levels of tyrosine hydroxylase (TH) were determined, using the Western blot technique that allows us to quantify and visualize TH enzyme changes caused by the MPTP and the effects of curcumin (CU). As shown in Figure 4, striatal TH expression was significantly decreased, by 43%, in offspring that were exposed to prenatal MPTP, as compared to the PBS control ( Figure 4, lane 1 vs 3). The figure also shows that exposure to postnatal 10 mg/kg of MPTP caused 16% reduction in TH expression ( Figure 4, lane 1 vs 2). Interestingly, MPTP has an additive effect on reducing TH, because, prenatal MPTP exposure coupled with postnatal exposure reduced TH by 68% ( Figure 4, lane 4, column 4). In offspring exposed to prenatal MPTP only, the reduction in TH was 43% ( Figure 4, lane 3) as compared to 16% in offspring exposed to the postnatal MPTP ( Figure 4, lane 2, column 2). It is of interest, also, that the prenatal exposure to CU partially reverses the changes in the expression of TH caused by the prenatal exposure to MPTP ( Figure 4, lane 3 vs 5), showing 18% improvement. Pre-exposure to CU also restores part of the losses in striatal TH caused by the paired exposure to prenatal-MPTP and postnatal-MPTP ( Figure 4, lane 4 vs 6, and highlighted in B, column 4 vs 6), showing a 50.3% improvement.
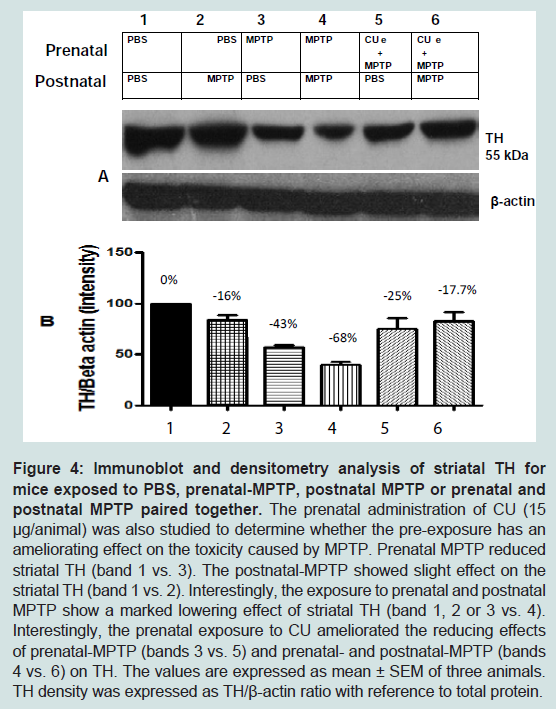
Figure 4: Immunoblot and densitometry analysis of striatal TH for mice exposed to PBS, prenatal-MPTP, postnatal MPTP or prenatal and postnatal MPTP paired together.The prenatal administration of CU (15 μg/animal) was also studied to determine whether the pre-exposure has an ameliorating effect on the toxicity caused by MPTP. Prenatal MPTP reduced striatal TH (band 1 vs. 3). The postnatal-MPTP showed slight effect on the striatal TH (band 1 vs. 2). Interestingly, the exposure to prenatal and postnatal MPTP show a marked lowering effect of striatal TH (band 1, 2 or 3 vs. 4). Interestingly, the prenatal exposure to CU ameliorated the reducing effects of prenatal-MPTP (bands 3 vs. 5) and prenatal- and postnatal-MPTP (bands 4 vs. 6) on TH. The values are expressed as mean ± SEM of three animals. TH density was expressed as TH/β-actin ratio with reference to total protein.
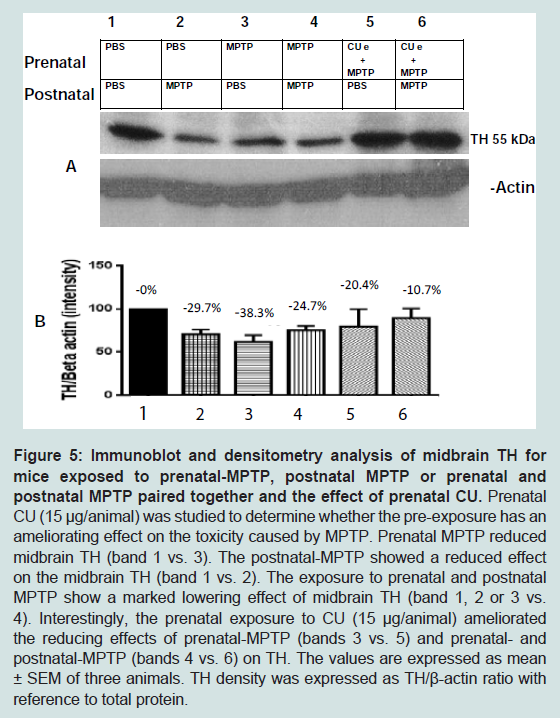
Figure 5: Immunoblot and densitometry analysis of midbrain TH for mice exposed to prenatal-MPTP, postnatal MPTP or prenatal and postnatal MPTP paired together and the effect of prenatal CU.Prenatal CU (15 μg/animal) was studied to determine whether the pre-exposure has an ameliorating effect on the toxicity caused by MPTP. Prenatal MPTP reduced midbrain TH (band 1 vs. 3). The postnatal-MPTP showed a reduced effect on the midbrain TH (band 1 vs. 2). The exposure to prenatal and postnatal MPTP show a marked lowering effect of midbrain TH (band 1, 2 or 3 vs. 4). Interestingly, the prenatal exposure to CU (15 μg/animal) ameliorated the reducing effects of prenatal-MPTP (bands 3 vs. 5) and prenatal- and postnatal-MPTP (bands 4 vs. 6) on TH. The values are expressed as mean ± SEM of three animals. TH density was expressed as TH/β-actin ratio with reference to total protein.
Discussion
The study confirms the possibility that neurodegenerative disorders, such as PD, may be caused by two phases of insults/affects: one occurring early in life, is without symptoms and sensitizes the neuronal system to changes that occur later in life. The changes that occur later in life could be caused by normal wear-and-tear of aging or by further insults to the neuronal system, that pairs with the early insults to cause threshold level harms that lead to neurodegeneration with accompanied functional changes.In this study, the prenatal or in-utero exposure to MPTP seems to be much more toxic than the exposure to the identical dosage of MPTP during adulthood. This is because prenatal exposure to 10 mg/kg of MPTP reduced DA by 52.8%, whereas the postnatal treatment of the same dosage of MPTP reduced DA by only 20%. A similar trend was also exhibited for TH, in which the prenatal exposure caused 43% reduction, and the postnatal exposure caused only 16% reduction. It should be noted also that neither prenatal MPTP, postnatal MPTP nor the pre-exposure to curcumin markedly affected the oxidation nor methylation of DA, based on the fact that the treatments that decreased/increased DA also proportionately decreased/increased DOPAC and 3-MT, the oxidative and methylated products of DA.
Prenatal exposure to MPTP also has a marked effect on motor activity, reducing total distance travelled (TD) to 42.4%, as compared to 25.1% following postnatal MPTP exposure. Moreover, prenatal exposure to MPTP may have a permanent effect, since the toxic effects of the in-utero exposure of pups to MPTP was evident in the adult mice. Interestingly, though the prenatal exposure to MPTP caused a reduction in birth weight of the pups but the effect lasted for 3-4 weeks. The recovery means that MPTP did not compromise postnatal development, evidence by the lack of changes in feeding and growth. More interestingly, CU was effective in blunting the reducing effect of prenatal MPTP on birth weight ( Figure 1), and CU reversed the reduction of motor activity caused by MPTP. Indeed, CU increased the total distance traveled by the mice, but it did not increase NM or MT. Curcumin also increased the speed of the animals, from an MPTPe-PBS value of 3.19 cm/sec to 6.89 cm/sec for the CUe-MPTPe-PBS animals and from a value of 3.09 cm/sec for the MPTPe-MPTP to 6.96 cm/sec for the CUe-MPTPe-MPTP group and to 6.9 cm/sec for the CUe-MPTPe-MPTP group.
Epidemiological studies have suggested that consumption of turmeric or CU by the Asian Indians is probably the reason for the low incidence of Alzheimer’s disease (AD) in East Indian, when compared to the Caucasians 34], so CU may also preserve the cholinergic neurons that are mostly responsible for the loss of memory seen in AD. A study exploring the effect of race and age on the prevalence of PD reported that there are approximately 40% less melanized nigral neurons in Indian brains when compared to Caucasian brains 35]. This is relevant, if melanization is caused by age-related changes, such as oxidative stress, for it was reported that there was no change in the number of nigral neurons with advancing age in the Asian Indian population 36]. Hence, the lower incidence of PD in India and lower numbers of melanized nigral neurons, suggest that there may be protective mechanisms for nigral neurons; probably associated with dietary habits. Interestingly, diet might have played an important role in protecting nigral neurons because chronic dietary consumption of turmeric, the source of CU, has been shown to offers neuroprotection in mouse models of PD 37-40].The neuroprotection that CU offers may involve its antioxidant 41,42], and anti-inflammatory [43-45] properties, as well as its ability to increase the density of dopaminergic neurons in the SN [46-48]. CU can penetrate the blood brain barrier, is relatively nontoxic, and shows few side effects at high doses 25,49]. An antioxidant role for CU would be complementary to the utility of rasagiline and selegiline in the therapy of PD, as these drugs serve as potent irreversible MAO B inhibitors [50], thus reducing DA oxidation. CU is known to be very safe, for when it is administered as standardized powder extract at a minimum of 95% purity, doses of up to 12 g per day showed no toxicity [51]. Interestingly, other natural products, such as carnosine [52], polyphenols constituents of fruits, vegetables, cereals, dry legumes, chocolate, tea, coffee and wine [53,54] as well as garlic [54] may all contribute to the prevention of degenerative diseases, such as PD.
MPTP was used in this study to target the NS DA neurons during their development in the mouse fetus. The goal was to cause subtle changes to the neuronal system that will make the dopaminergic neurons susceptible to toxic changes that occur later in life. For this study, we also used the postnatal administration of MPTP to mimic the stress that the NS DA neurons may encounter during adulthood and aging. Together the two courses of MPTP allow us to evaluate the effects of prenatal and postnatal assaults on the NS DA system. CU was administered prenatally to test its effectiveness in blocking the changes caused by MPTP by comparing the MPTPe-PBS group with CUe-MPTPe-PBS group. The results clearly show that CU reduced or prevented the toxicities caused by prenatal MPTP. CU restores the loss of DA caused by MPTP, from the prenatal MPTP loss of 52.8% to only 2.7% loss when CU was given. CU also reversed the reduction in motor activity caused by MPTP. The midbrain contains the substantia nigra, the location for the cell-bodies of the dopamine neurons that project to the neostriatum. Interestingly, the levels of TH that occur in the midbrain of MPTP exposed offspring treated with CU, suggests that the transport of TH to the striatum from the midbrain kept up with the level of the changes in midbrain TH caused by CU.
Conclusion
The results demonstrate that at least two phases may be involved in causing PD-like impairments: one occurs early in life, and the other later in life and involves changes caused by aging or by other harmful incidences. This study shows that CU can blunt changes that occur during both phases. The objective of the study was to test the hypothesis that two stages of impairment are involved in neurological disorders, such as PD. To prove the first stage concept, MPTP was administered to pregnant mice during the period of differentiation of the basal ganglia dopamine neurons. For the second stage, the prenatal MPTP-exposed offspring were challenged with 10 mg/kg MPTP, to determine if such a low dose of MPTP will affect the NS DA system in the offspring that were exposed to MPTP in utero. The data clearly show that the prenatal exposure to MPTP sensitizes the NS DA system, on the basis that the postnatal administration of MPTP caused dramatic toxicity in the prenatal-MPTP exposed group. Postnatal MPTP reduced TH by 68% and DA by 67.2% in offspring that were exposed to prenatal MPTP. The same MPTP postnatal treatment reduced TH by only 16% and DA by 20% in the controls offspring that were exposed to prenatal PBS. Thus, the MPTP challenge mimics, in a short period of time, the age related and protracted changes that cause PD in the adult human. Furthermore, we studied offspring that were exposed to prenatal MPTP (1st phase insult) as well as animals that were exposed to both prenatal and postnatal MPTP (both phases of insults) to determine if prenatal CU can ameliorate the toxic effects of MPTP. However, no literature is available illustrating the efficacy of CU in early and late phases of Parkinson’s disease. So this study is the first reported outcome showing that CU prevented the MPTP induced reduction of DA and TH, restored the oxidative and methylated turnover rate for DA and increased motor activity. Therefore, the study demonstrates that prenatal CU treatment blocked MPTP effects in the striatum and midbrain, signifying that curcumin may serve as a neuroprotective agent for PD.Acknowledgement
The works were supported by NIHRO1NS041674 and NIHR21NS049623. The authors wish to acknowledge the financial support from Dr. Bernard Crowell, Pinnacle Orthopedics. P.A., 13100 Chenel Parkway, Little Rock AR 72223.References
- Dauer W, Pizeborski S (2003) Parkinson’s disease: mechanisms and models. Neuron 39: 889-909.
- Nagatsu I, Kondo Y, Inagaki S, Kojima H, Nagatsu T (1979) Immunofluorescent and biochemical studies on tyrosine hydroxylase and dopamine-beta-hydroxylase of the bullfrog sciatic nerves. Histochemistry 18: 103-109.
- Heikkila RE, Sonsalla PK (1992) The MPTP treated mouse as a model of parkinsonism: how good is it? Neurochem Int 20: 299S-303S.
- Beal MF (2000) Oxidative metabolism. Ann N Y Acad Sci 924: 164-169.
- Przedborski S (2005) Pathogenesis of nigral cell death in Parkinson’s disease. Parkinsonism Relat Disord 11: Suppl 1: S3-S7.
- Jha N, Jurma O, Lalli G, Liu Y, Pettus EH, et al. (2000) Glutathione depletion in PC12 results in selective inhibition of mitochondrial complex 1 activity: implications for Parkinson’s disease. J Biol Chem 275: 26096-26101.
- Kühnau J (1976) The flavonoids. A class of semi- essential food components: their role in human nutrition. World Rev Nutr Diet 24: 117-191.
- Hertog MG, Feskens EJ, Hollman PC, Katan MB, Kromhout D (1993) Dietary antioxidant flavonoids and risk of coronary heart disease: the Zutphen Elderly Study. Lancet 342: 1007-1011.
- Vinson JA, Jang J, Dabbagh JA, Serry MM (1995) Plant flavonoids especially tea flavonoids are powerful antioxidants using an in vitro oxidation model for heart disease. J Agric Food Chem 43: 2800-2802.
- Wiseman SA, Balentine DA, Frei B (1997) Antioxidants in tea. Crit Rev Food Sci Nutr 37: 705-718.
- Saija A, Scalese M, Lanza M, Marzullo D, Bonia F, et al. (1995) Flavonoids as antioxidant agents: importance of their interaction with biomembranes. Free Radic Biol Med 19: 481-486.
- Middleton E Jr, Kandaswami C, Theoharides TC (2000) The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol Rev 52: 673-751.
- Sudheesh S, Sandhya C, Sarah Koshy A, Vijayalakshmi NR (1999) Antioxidant activity of flavonoids from Solanum melongena. Phytother Res 13: 393-396.
- Park YC, Pae HO, Yoo JC, Choi BM, Jue DM, et al. (1999) Chloroquine inhibits inducible nitric oxide synthase expression in murine peritoneal macrophages. Pharmacol Toxicol 85: 188-191.
- Ishiwa J, Sato T, Mimaki Y, Sashida Y, Yano M, et al. (2000) A citrus flavonoid, nobiletin, suppresses production and gene expression of matrix metalloproteinase 9/gelatinase B in rabbit synovial fibroblast. J Rheumatol 27: 20-25.
- Matsuoka Y, Hasegawa H, Okuda S, Muraki T, Uruno T, et al. (1995) Ameliorative effects of tea catechins on active oxygen-related nerve cell injuries. J Pharmacol Exp Ther 274: 602-608.
- Pelzer LE, Guardia T, Osvaldo Juarez A, Guerreiro E (1998) Acute and chronic antiinflammatory effects of plant flavonoids. Farmaco 53: 421-424.
- Di Carlo G, Mascolo N, Izzo AA, Capasso F (1999) Flavonoids: old and new aspects of a class of natural therapeutic drugs. Life Sci 65: 337-353.
- Murakami A, Nakamura Y, Ohto Y, Yano M, Koshiba T, et al. (2000) Suppressive effects of citrus fruits on free radical generation and nobiletin, an anti-inflammatory polymehhoxyflavonoid. Biofactors 12: 187-192.
- Hoult JR, Moroney MA, Payá M (1994) Actions of flavonoids and coumarins on lipoxygenase and cyclooxygenase. Methods Enzymol 234: 443-454.
- Lodha R, Bagga A (2000) Traditional Indian systems of medicine. Ann Acad Med Singapore 29: 37-41.
- Srimal RC, Dhawan BN (1973) Pharmacology of diferuloyl methane (curcumin), a non-steroidal anti-inflammatory agent. J Pharm Pharmacol 25: 447-452.
- Tobinai K, Shimoyama M, Inoue S, Takayasu S, Mikuni C, et al. (1992) Phase I study of YK -176 (2’deoxycoformycin) in patient with adult T- cell Leukemia- lymphoma. The DCF Study Group. Jpn J Clin Oncol 22: 164-171.
- Uozumi K, Hanada S, Ohno N, Ishitsuka K, Shimotakahara S, et al. (1995) Combination chemotherapy (RCM protocol: response-oriented cyclic multidrug protocol) for the acute or lymphoma type adult T-cell leukemia. Leuk Lymphoma 18: 317- 323.
- Lim GP, Chu T, Yang F, Beech W, Frautschy SA, et al. (2001) The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J Neurosci 21: 8370-8371.
- Yang F, Lim GP, Begum AN, Ubeda OJ, Simmons MR, et al. (2005) Curcumin inhibits formation of amyloid beta oligomers and fibril binds plaques and reduces amyloid in vivo. J Biol Chem 280: 5892-5901.
- Chen C, Johnston TD, Wu G, Ranjan D (2006) Curcumin has potent liver preservation properties in an isolated perfusion model. Transplantation 82: 931-937.
- Qin XY, Cheng Y, Yu LC (2009) Potential protection of curcumin against intracellular amyloid beta-induced toxicity in cultured rat prefrontal cortical neurons. Neurosci Lett 480: 21-24.
- Betarbet R, Sherer TB, Greenamyre JT (2002) Animal models of Parkinson’s disease. Bioessays 24: 308-318.
- Grunblatt E, Mandel S, Youdim MB (2000) MPTP and 6-hydroxydopamine-induced neurodegeneration as models for Parkinson’s disease: neuroprotective strategies. J Neurol 247 Suppl 2: II95-II102.
- Muthian G, Mackey V, King J, Charlton CG (2010) Modeling a sensitization stage and a precipitation stage for Parkinson's disease using prenatal and postnatal 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine administration. Neuroscience 169: 1085-1093.
- Muthian G, King J, Dent L, Smith M, Mackey V, et al. (2012) Prenatal and postnatal exposures to 1-Methyl-4-phenyl-1,2,3,6-tetra hydropyridine (MPTP) impaired mouse midbrain dopamine system and may produce a predisposing and inducing model for Parkinson’s disease. J Behav Brain Sci 2: 485-494.
- Nicotra A, Parvez SH (2000) Cell death induced by MPTP, a substrate for monoamine oxidase B. Toxicology 153: 157-166.
- Ganguli M, Chandra V, Kamboh MI (2000) Apolipoprotein E polymorphism and Alzheimer disease: The Indo-US Cross -National Dementia study. Arch Neurol 57: 824-830.
- Muthane U, Yasha TC, Shankar SK (1998) Low numbers and no loss of melanized nigral neurons with increasing age in normal human brains from India. Ann Neurol 43: 283-287.
- Alladi PA, Mahadevan A, Yasha TC, Raju TR, Shankar SK, et al. (2009) Absence of age-related changes in nigral dopaminergic neurons of Asian Indians: relevance to lower incidence of Parkinson’s disease. Neuroscience 159: 236-245.
- Mythri RB, Harish G, Dubey SK, Misra K, Bharath MM (2011) Glutamoyl diester of the dietary polyphenol curcumin offers improved protection against peroxynitrite-mediated nitrosative stress and damage of brain mitochondria in vitro: implications for Parkinson’s disease. Mol Cell Biochem 347: 135-143.
- Vajragupta O, Boonchoong P, Watanabe H, Tohda M, Kummasud N, et al. (2003) Manganese complexes of curcumin and its derivatives: evaluation for the radical scavenging ability and neuroprotective activity. Free Radic Biol Med 35: 1632-1644.
- Zbarsky V, Datla KP, Parkar S, Rai DK, Aruoma OI, et al. (2005) Neuroprotective properties of the natural phenolic antioxidants curcumin and naringenin but not quercetin and fisetin in a 6-OHDA model of Parkinson’s disease. Free Radic Res 39: 1119-1125.
- Rajeswari A (2006) Curcumin protects mouse brain from oxidative stress caused by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Eur Rev Med Pharmacol Sci 10: 157-161.
- Zbarsky V, Datla KP, Parkar S, Rai DK, Aruoma OI, et al. (2005) Neuroprotective properties of the natural phenolic antioxidants curcumin and narigenin but not quercetin and fisetin in a 6-OHDA model of Parkinson’s disease. Free Radic Res 39: 1119-1125.
- Zhao BL, Li XJ, He RG, Cheng SJ, Xin WJ (1989) Scavenging effect of extracts of green tea and natural antioxidants on active oxygen radicals. Cell Biophys 14: 175-185.
- Xu YX, Pindolia KR, Janakiraman N, Chapman RA, Gautam SC (1998) Curcumin inhibits 1L-1 alpha and TNF-alpha induction of AP-1 and NF-kB DNA-binding activity in bone marrow stromal cells. Hematopathol Mol Hematol 11: 49-62.
- Chan MM, Huang HI, Fenton MR, Fong D (1998) In vivo inhibition of nitric oxide synthase gene expression by curcumin, a cancer preventive natural product with anti-inflammatory properties. Biochem Pharmacol 55: 1955-1962.
- Zhang F, Altorki NK, Mestre JR, Subbaramaiah K, Dannenberg AJ (1999) Curcumin inhibits cyclooxygenase-2 transcription in bile acid- and phorbol ester-treated human gastrointestinal epithelial cells. Carcinogenesis 20: 445- 451.
- Vajragupta O, Boonchoong P, Watanabe H, Tohda M, Kummasud N, et al. (2003) Manganese complexes of curcumin and its derivatives: evaluation for the radical scavenging ability and neuroprotective activity. Free Radic Biol Med 12: 1632-1644.
- Zbarsky V, Datla KP, Parkar S, Rai DK, Aruoma OI, et al. (2005) Neuroprotective properties of the natural phenolic antioxidants curcumin and naringenin but not quercetin and fisetin in a 6-OHDA model of Parkinson's disease. Free Radic Res 39: 1119-1125.
- Rajeswari A (2006) Curcumin protects mouse brain from oxidative stress caused by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Eur Rev Med Pharmacol Sci 10: 157-161.
- Sharma RA, Euden SA, Platton SL, Cooke DN, Shafayat A, et al. (2004) Phase I clinical trial of oral curcumin: biomarkers of systemic activity and compliance. Clin Cancer Res 15: 6847-6854.
- Borstnar R, Repic M, Krzan M, Mavri J, Vianello R (2011) Irreversible inhibition of monoamine oxidase B by the antiparkinsonian medicines rasagiline and selegiline: A computational study. Eur J Org Chem 2011: 6419-6433.
- Lao CD, Ruffin MT 4th, Normolle D, Heath DD, Murray SI, et al. (2006) Dose escalation of curcuminoid formulation. BMC Complement Altern Med 6: 10.
- Hipkiss AR (2014) Aging risk factors and Parkinson’s disease: contrasting roles of common dietary constituents. Neurobiol Aging 35: 1469-1472.
- Scalbert A, Manach C, Morand C, Remesy C, Jimenez L (2005) Dietary polyphenols and the prevention of diseases. Crit Rev Food Sci Nutr 45: 287-306.
- Pavlin M, Repič M, Vianello R, Mavri J (2015) The chemistry of neurodegeneration: Kinetic data and their implications. Mol Neurobiol [Epub ahead of print].